
Содержание
Пути воздействия организмов на среду обитания
1. Пути воздействия организмов на среду обитания
Живые организмы сильно влияют на среду
обитания уже тем, что живут в ней. Они
дышат, питаются, выделяют продукты
обмена, растут и размножаются,
перемещаются в пространстве, проявляют
разные формы активности. В
результате этого изменяются и
газовый состав воздуха, и
микроклимат, и почва, и
чистота вод, и другие
особенности местообитаний.
И хотя воздействие
каждого отдельного
организма на
окружающую среду
может быть мало,
масштабы суммарной
активности живых
существ огромны.
Влияние организмов на
среду обитания
называют их
средообразующей
деятельностью.
4. Влияние растений на климат и водный режим.
Фотосинтез —
главный источник
кислорода в земной
атмосфере. Растения
создают условия для
дыхания миллиардам
живых существ,
включая людей.
Потребности в
кислороде лишь одного
человека за 70—80 лет
жизни составляют
несколько десятков
тонн.
Если представить, что
фотосинтез на планете
прекратится, весь
кислород атмосферы
израсходуется всего за
2000 лет. Содержание в
воздухе азота,
углекислого газа и
ряда других соединений
также зависит от
жизнедеятельности
различных организмов.
Поглощение и испарение
воды наземными
растениями влияет на
водный режим их
местообитаний и на
климат в целом. За час
выделяется до 2,5 г воды с
каждого квадратного
дециметра листвы. Это
составляет ежечасно
многие тонны воды с
гектара. Одно только
дерево березы испаряет в
день до 100 л воды.
Увлажняя воздух,
задерживая движение
ветра, растительность
создает особый
микроклимат,
смягчающий условия
существования многих
видов. В лесу
колебания температуры
в течение года и суток
меньше, чем на
открытых
пространствах.
Леса сильно изменяют
также условия влажности:
снижают уровень
грунтовых вод,
задерживают осадки,
способствуют осаждению
росы и тумана,
предотвращают эрозию
почвы. В них возникает
В них возникает
особый световой режим,
позволяющий
тенелюбивым видам расти
под пологом более
светолюбивых.
9. Почвообразующая деятельность живых организмов.
Совместная
деятельность
множества организмов
создает почву.
Сбрасывая ежегодно
листву, растительность
образует на
поверхности земли
слой мертвого
органического
вещества.
Этот слой растительного
опада служит источником
пищи и средой обитания для
огромного количества
мелких организмов —
бактерий, грибов,
животных, которые
разрушают и
перерабатывают его до
неорганических молекул.
Освободившиеся
минеральные вещества
вновь идут на питание
растений. Некоторая часть
органических веществ
превращается в почвенный
гумус.
Это сложные соединения,
которые улучшают
структуру почвы, ее влагои воздухопроницаемость.
Тем самым улучшаются
условия для развития
корней растений. Таким
образом, процесс
образования почвы в первую
очередь зависит от пищевой
активности множества живых
существ, использующих
энергию мертвого
органического вещества.
Каждый комочек почвы
содержит миллионы
клеток различных
микроорганизмов. Кроме
них, на каждый
квадратный метр почвы
приходятся сотни тысяч
мелких животных,
различимых только в
микроскоп, и тысячи —
видимых простым глазом.
Особенно важна для
жизни почвы
деятельность дождевых
червей.
Их нормальная
численность в лесах и на
лугах составляет от
нескольких десятков до
нескольких сотен особей
на квадратный метр.
Дождевые черви
разрыхляют и
перемешивают слои
почвы, улучшают условия
для прорастания корней
растений, затягивают
вглубь растительные
остатки. Выделения из их
кишечников представляют
прочные органоминеральные комочки.
Большое их количество в почве резко
улучшает ее структуру и повышает
плодородие. При высокой численности
дождевые черви за год могут образовывать
до 120 т таких комочков на 1 га. Таким
образом, почва — это среда
обитания, созданная
деятельностью самих
живых организмов.
Деятельность животных
может иногда определять
особенности ландшафта.
Настоящие плотины
устраивают бобры.
Крупные животные норники, такие, как
суслики или сурки,
обеспечивают мозаичность
растительного и
почвенного покрова, так
как за счет выбросов
почвы формируется
микрорельеф,
перераспределяющий
осадки и видовой состав
растений.
16. Влияние водных организмов на качество природных вод.
Качество воды в
водоемах во многом
зависит от фильтрующих
животных. Многие из
них ведут сидячий образ
жизни или «парят» в
толще воды, отцеживая
из окружающей среды
пищевые частицы.
Многочисленные
пластинчатожаберные
моллюски, такие, как
устрицы и мидии в морях,
• а в пресных водах —
перловицы, беззубки,
ресничками на ротовых
лопастях подгоняют
воду к ротовому
отверстию и сортируют
взвесь. При этом
частицы, непригодные в
пищу, формируются в
комочки и осаждаются
на дно.
Мелкие рачки, такие,
как дафнии,
отцеживают пищевую
взвесь густыми
щеточками щетинок на
своих конечностях.
Личинки мошек в
ручьях
отфильтровывают
пищу пучками щетинок
на голове, а личинки
комаров — щетками на
верхней губе.
Активно
процеживают воду
через жаберный
аппарат некоторые
рыбы, как
например
толстолобик и
китовая акула.
Фильтрационное питание
наблюдается у 40 тысяч
видов водных животных.
В результате этой
деятельности происходит
биологическое
самоочищение водоемов, и
от него зависит качество
воды. Одна перловица
длиной 5—6 см при
температуре 20 °С очищает
до 16 л воды в сутки. В
прудах и озерах, где много
мелких рачков, весь объем
воды пропускается через их
фильтровальный аппарат
всего за один день.
Один квадратный метр
морского мелководья,
густо заселенный
моллюсками мидиями,
за сутки может очистить
до 280 м 3 воды. Таким
Таким
образом, чистота и
прозрачность
природных вод —
результат деятельности
живых организмов.
Способность
организмов изменять
среду обитания широко
используется в
хозяйственной практике.
Для улучшения
микроклимата, условий
увлажнения и защиты
полей от иссушающих
ветров в степных районах
сажают лесополосы, для
очистки воздуха в
городах и курортных
зонах создают парки и
сады.
На водоочистительных
станциях строят
специальные емкости, где
поддерживается высокая
активность мелких
фильтраторов. Используя
почвообразующую
деятельность животных и
микроорганизмов,
предприятия по переработке
органических отходов
производят удобрения для
внесения в истощенные
почвы.
Условия жизни людей
на Земле зависят от
средообразующей роли
миллиардов живых
организмов. И состав
воздуха, и качество вод,
и почвенное
плодородие, и
микроклимат
складываются из их
суммарной
деятельности.
25. Примеры и дополнительная информация
1. Одна из новейших технологий
переработки отходов свиноводческих ферм,
которые сильно загрязняют окружающую
среду, основана на использовании
средоопреобразующей деятельности личинок
комнатных мух. В специальных контейнерах
навоз засевают яйцами мух. Личинки,
вышедшие из яиц, питаются им и, кроме
того, выделяют наружу вещества,
убивающие болезнетворных бактерий и яйца
паразитов.
Через 5 суток субстрат полностью
перерабатывается личинками в рыхлый
стерильный перегной, очень ценный как
органическое удобрение. Выросших личинок
извлекают из субстрата автоматизированными
методами и используют как белковый корм
на птицефермах и в рыбных хозяйствах.
Таким образом, вид, который ранее считался
только вредным, превращен в хозяйственно
ценный.
2. Для очистки промышленных и городских
сточных вод от органических веществ
используют деятельность бактерий и мелких
фильтраторов (инфузорий, коловраток и др.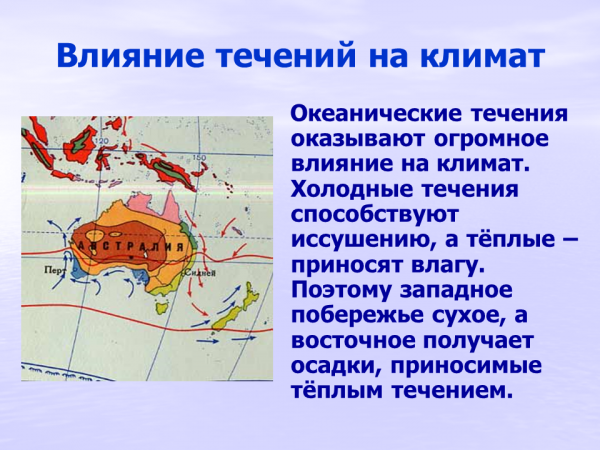 ).
).
Один из видов очистных сооружений —
аэротенки. Это длинные емкости глубиной
5 м и шириной 10 м, через которые
протекает сточная жидкость. Со дна
аэротенка подается воздух в виде мелких
пузырьков, поднимающихся кверху.
Ток воздуха создает благоприятные
кислородные условия для микроорганизмов
и простейших, которые размножаются в
огромном количестве. Они очищают воду,
образуя хлопья так называемого
«активного ила». Из аэротенков вода
поступает в отстойники, где «активный
ил» оседает на дно, а затем вновь
используется для зарядки аэротенка.
3. Зеленые насаждения в
городе намного
улучшают микроклимат.
В городских парках в
жаркий день температура
на 6—8° ниже, чем на
улицах. Даже возле
газонов на 2—3°
прохладнее, чем на
тротуаре, за счет
испарения растениями
влаги. Заметно
изменяется и состав
городского воздуха.
Одно дерево выделяет
столько кислорода,
сколько нужно для
дыхания 4 человек.
Кроме того, растения
поглощают примеси
некоторых ядовитых
газов и выделяют
летучие вещества —
фитонциды, которые
губительны для бактерий,
содержащихся в воздухе.
Один гектар парка из
лиственных деревьев
задерживает за год до
100 т пыли. В городах с
интенсивной
промышленностью
рекомендуют высаживать
особо газоустойчивые
деревья и кустарники:
различные тополя, тую
западную, клен
американский, черемуху,
бузину красную и др.
4. Подсчитано, что в
Волгоградском
водохранилище мелкие
двустворчатые
моллюски дрейссены
с апреля по ноябрь
отфильтровывают 840
млрд м 3 воды, что в
24 раза превышает
полный объем
водохранилища. При
этом ими осаждается
на грунт 29 млн т
несъедобной взвеси, в
среднем более 8 кг на
каждый квадратный
метр.
5. Среднее число нор
млекопитающих на 1 га
составляет в лиственных
лесах около 1000, в
лесостепи — 7500, в
степи — 5000, в
пустынях — 1500.
Ежегодно норы
подновляются или
роются на новом месте.
Перерытые участки
заселяются сорными
растениями, которые
способны прорастать
только на нарушенных
местах.
Эти растения, которые
широко распространены
в настоящее время на
пахотных почвах,
существовали задолго
до появления
земледелия и обязаны
своим происхождением
деятельности роющих
животных.
Климат. Атмосферный климат
Важной частью общих представлений В.В. Докучаева о почве является учение о факторах почвообразования. В.В. Докучаев установил, что почва есть продукт совокупной деятельности горных пород, климата, растительных и животных организмов, рельефа местности и возраста страны (времени). Для того чтобы изучать почву — функцию почвообразователей, необходимо исследовать и сами почвообразователи. Это положение легло в основу важнейших разделов почвоведения — генезиса и географии почв. Знание сущности и истории влияния природных факторов на формирование почв позволяет предсказать тип почвенного покрова на территории, которая еще не изучена.
Климат — один из важнейших факторов почвообразования и географического распространения почв — в значительной степени определяется космическими причинами (количеством энергии, получаемой земной поверхностью от Солнца). Климат влияет на почвообразование непосредственно, определяя энергетический уровень и гидротермический режим почв, и косвенно через другие факторы почвообразования (растительность, почвообразующую породу, рельеф и др.).
Непосредственная роль климата проявляется в том, что под его действием формируются тепловой и водный режимы почв, которые оказывают влияние на характер и интенсивность всех физических, химических и биологических процессов, протекающих в почвах. С климатическими условиями тесно связаны процессы превращения минеральных соединений в почве (направление и темпы выветривания, миграция и аккумуляция продуктов почвообразования и др.), процессы ветровой и водной эрозии. Определенные сочетания температуры и увлажнения обусловливают тип растительности, темпы создания и разрушения органического вещества, состав и интенсивность деятельности почвенной микрофлоры и фауны. С климатическими условиями связана энергетика почвообразования. Климат играет важнейшую роль в закономерном размещении почв на Земле.
С климатическими условиями связана энергетика почвообразования. Климат играет важнейшую роль в закономерном размещении почв на Земле.
Главный источник энергии для почвенных процессов — солнечная радиация, а основной источник увлажнения — атмосферные осадки.
Среднегодовая температура воздуха на территории Российской Федерации возрастает с севера на юг и с востока на запад. Увеличение температуры воздуха с севера на юг происходит за счет повышения летних температур. В соответствии с изменением температуры воздуха меняется и обеспеченность теплом природных поясов. Границы поясов характеризуются определенными суммами среднесуточных температур выше 10°С. Такие температуры называются активными, поскольку это основной период вегетации растений. На крайнем севере Российской Федерации период со средней суточной температурой выше 10°С отсутствует. Суммы температур воздуха за период с устойчивой температурой выше 10°С меняются на материке от 400–600° и менее на севере до 3200 и более на юге. Растет и число дней в году с активными температурами воздуха.
Растет и число дней в году с активными температурами воздуха.
Приход солнечной энергии на земную поверхность подчиняется ритмам различной продолжительности (суточным, сезонным, годовым, многолетним). Самым теплым месяцем является июль (в ряде регионов Дальнего Востока — август), самым холодным — январь. В то время как температуры июля на территории Российской Федерации нарастают с севера на юг и мало различаются с запада на восток, изолинии зимних температур воздуха имеют меридиональное простирание, и температуры января существенно снижаются к востоку, определяя степень континентальности климата.
Континентальность климата вычисляется по формуле, предложенной Н.Н. Ивановым, К = A × 100% / 0.33ϕ, где А — годовая амплитуда температуры из среднемесячных ее величин, ϕ — широта местности.
Таежно-лесная зона Русской равнины имеет умеренно континентальный климат, степная — среднеконтинентальный. Почти вся Западно-Сибирская равнина, за исключением полуостровов Ямал и Гыдан и крайнего юга, лежит в области среднеконтинентального климата. Средняя и Восточная Сибирь характеризуется очень континентальным и резко континентальным климатом. Степень континентальности вновь снижается на Тихоокеанском побережье. И лишь Кольский полуостров, побережье Баренцева моря и западное побережье Новой Земли отличаются океаническим климатом.
Средняя и Восточная Сибирь характеризуется очень континентальным и резко континентальным климатом. Степень континентальности вновь снижается на Тихоокеанском побережье. И лишь Кольский полуостров, побережье Баренцева моря и западное побережье Новой Земли отличаются океаническим климатом.
Вечная мерзлота — это мерзлые толщи горных пород, не оттаивающие в течение длительного времени, от нескольких до десятков и сотен тысяч лет. Верхние горизонты мерзлых пород (от 0,5 до 3–5 м) ежегодно (сезонно) протаивают и промерзают, а глубже залегает слой постоянно мерзлого грунта, который и называют вечной, или многолетней, мерзлотой. Вечная мерзлота занимает почти 2/3 территории России (Карта Вечная мерзлота составлена с использованием опубликованных материалов [33]).
Годовая сумма осадков на территории РФ колеблется от значений более 1450 мм до 250 мм и менее. Максимум наблюдается в южных горных системах (Кавказ, Алтай, Саяны, Сихотэ-Алинь), минимум — на азиатском побережье и островах Ледовитого океана, Центральноякутской, Индигирской и Колымской низменностях, а также в Прикаспийской полупустыне. В пределах почвенных зон годовое количество осадков уменьшается с запада на восток, а в Приморье с востока на запад с муссонным характером их выпадения (количество осадков теплого полугодия в 4–8 раз больше, чем холодного).
В пределах почвенных зон годовое количество осадков уменьшается с запада на восток, а в Приморье с востока на запад с муссонным характером их выпадения (количество осадков теплого полугодия в 4–8 раз больше, чем холодного).
Разность осадков и испаряемости отражает влагообеспеченность климата. На Русской и ЗападноСибирской равнинах с севера до южных границ зоны дерново-подзолистых почв количество осадков превышает испаряемость. Южнее, а также на территории Средней и Восточной Сибири возможное испарение больше суммы осадков.
Атмосферный климат, преломляясь в почве, формирует климат почвы. Климат почвы — это водный, воздушный и температурный режимы в многолетнем цикле.
И.С. Урусевская
Иллюстрации:
- Суммы температур воздуха за период с устойчивой температурой выше 10°, масштаб 1:40 000 000
- Число дней в году со средней суточной температурой воздуха выше 10°, масштаб 1:40 000 000
- Длительность безморозного периода, масштаб 1:40 000 000
- Температура воздуха.
 Январь, масштаб 1:40 000 000
Январь, масштаб 1:40 000 000 - Температура воздуха. Июль, масштаб 1:40 000 000
- Континентальность климата, масштаб 1:60 000 000
- Снежный покров, масштаб 1:40 000 000
- Вечная мерзлота, масштаб 1:40 000 000
- Количество осадков. Год, масштаб 1:40 000 000
- Количество осадков. Теплый период, масштаб 1:40 000 000
- Разность осадков и испаряемости, масштаб 1:40 000 000
 Январь, масштаб 1:40 000 000
Январь, масштаб 1:40 000 000← Назад
На уровень выше
Далее →
Влияние режимов влажности почвы на рост, фотосинтетическую способность, биохимию листа и репродуктивную способность инвазивных сельскохозяйственных сорняков; Lactuca serriola
1. Джалил К.А., Маниваннан П., Вахид А., Фарук М., Аль-Джубури Х.Дж., Сомасундарам Р. и соавт.
Стресс засухи у растений: обзор морфологических характеристик и состава пигментов. Int J Agric Biol. 2009 г.; 11(1): 100–105. [Google Scholar]
2. Чавес М.М., Мароко Дж.П., Перейра Дж.С. Понимание реакции растений на засуху — от генов до всего растения. Функция биологии растений. 2003 г.; 30(3): 239–264. [Google Scholar]
Функция биологии растений. 2003 г.; 30(3): 239–264. [Google Scholar]
3. Bohnert HJ, Gong Q, Li P, Ma S. Разгадка механизмов устойчивости к абиотическим стрессам — запуск геномики. Curr Opin Plant Biol. 2006 г.; 9(2): 180–188. 10.1016/j.pbi.2006.01.003
[PubMed] [CrossRef] [Google Scholar]
4. Hu Y, Schmidhalter U. Засуха и засоление: сравнение их влияния на минеральное питание растений. J Plant Nutr Soil Sci. 2005 г.; 168(4): 541–549. [Google Scholar]
5. Паттерсон Д.Т. Влияние экологического стресса на взаимодействие сорняков и сельскохозяйственных культур. Сорняк наук. 1995; 43: 483–490. [Google Scholar]
6. Зимдал Р.Л. Основы науки о сорняках; Третье издание
Калифорния, Лондон: Academic Press; 2018. [Google Scholar]
7. Баджва А.А., Махаджан Г., Чаухан Б.С. Нетрадиционные стратегии борьбы с сорняками в современном сельском хозяйстве. Сорняк наук. 2015 г.; 63(4): 723–747. [Google Scholar]
8. Chaves MM, Pereira JS, Maroco J, Rodrigues ML, Ricardo CPP, OsÓRio ML, et al.
Как растения справляются с водным стрессом в полевых условиях?
Фотосинтез и рост. Энн Бот. 2002 г.; 89(7): 907–916. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Bosco de Oliveira A, Alencar NLM, Gomes-Filho E. Сравнение воздействия водного и солевого стресса на рост и развитие растений В: Akıncı S, редактор. Реакции организмов на водный стресс: сельскохозяйственные и биологические науки. Издатель IntechOpen; 2013. [Google Scholar]
10. Осакабе Ю., Осакабе К., Шинозаки К., Тран Л-СП. Реакция растений на водный стресс. Фронт завод науч. 2014; 5: 86
10.3389/fpls.2014.00086
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11. Фарук М., Вахид А., Кобаяши Н., Фудзита Д., Басра СМА. Стресс растений от засухи: последствия, механизмы и управление В: Lichtfouse E, Navarrete M, Debake P, Véronique S, Alberola C, редакторы. Устойчивое сельское хозяйство. Дордрехт: Springer, Нидерланды; 2009. С. 153–188. [Google Scholar]
12. Taiz L, Zeiger E. Физиология растений; Пятое издание
Физиология растений; Пятое издание
Sinauer Associates: Сандерленд; 2002. [Google Scholar]
13. Мэнсфилд Т., Аткинсон С. Поведение устьиц у растений, подвергающихся водному стрессу. Биол растений (США). 1990. [Google Scholar]
14. Анджум Ф., Ясин М., Расул Э., Вахид А., Анджум С. Водный стресс у ячменя ( Hordeum vulgare L.). II. Влияние на химический состав и содержание хлорофилла. Пакистан J Agric Sci. 2003 г.; 40: 45–49. [Google Scholar]
15. Fu J, Huang B. Участие антиоксидантов и перекисного окисления липидов в адаптации двух прохладных трав к локальному стрессу от засухи. Бот Environment Exp. 2001 г.; 45(2): 105–114. [PubMed] [Google Scholar]
16. Монахова О.Ф., Чернядьев И.И. Защитная роль Картолина-4 у растений пшеницы при воздействии почвенной засухи. Приложение Biochem Microbiol. 2002 г.; 38(4): 373–380. [Академия Google]
17. Бартелс Д., Сункар Р. Засухо- и солеустойчивость растений. Crit Rev Plant Sci. 2005 г.; 24(1): 23–58. [Google Scholar]
18. Секи М., Умэдзава Т., Урано К., Шинозаки К. Регуляторные метаболические сети в реакциях на засуху. Curr Opin Plant Biol. 2007 г.; 10(3): 296–302. 10.1016/j.pbi.2007.04.014
Секи М., Умэдзава Т., Урано К., Шинозаки К. Регуляторные метаболические сети в реакциях на засуху. Curr Opin Plant Biol. 2007 г.; 10(3): 296–302. 10.1016/j.pbi.2007.04.014
[PubMed] [CrossRef] [Google Scholar]
19. Донохью К. Завершение цикла: материнские эффекты как недостающее звено в истории жизни растений. Философские труды Королевского общества B: Биологические науки. 2009 г.; 364 (1520): 1059–1074. 10.1098/рстб.2008.0291
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
20. Jairus R, Dennis P, Sebastian R, Mark B, Patrick FE. Вызванные затенением и засухой изменения содержания фенолов в семенах дикого овса ( Avena fatu a L.). Журнал физиологии стресса и
Биохимия. 2010 г.; 6(4). [Google Scholar]
21. Steadman KJ, Ellery AJ, Chapman R, Moore A, Turner NC. Температура созревания и осадки влияют на характеристики покоя семян райграса однолетнего ( Лолиум жесткий ). Aust J Agric Res. 2004 г.; 55 (10): 1047–1057. [Google Scholar]
22. Смит Дж., Гриффитс Х. Реакция растений на дефицит воды — от клетки к сообществу. J Опытный бот. 1992 год; 43(250): 29–37. [Google Scholar]
Смит Дж., Гриффитс Х. Реакция растений на дефицит воды — от клетки к сообществу. J Опытный бот. 1992 год; 43(250): 29–37. [Google Scholar]
23. D’Andrea L, Broennimann O, Kozlowski G, Guisan A, Morin X, Keller-Senften J, et al.
Изменение климата, антропогенное воздействие и расширение ареала Lactuca serriola (Asteraceae) на север. J Биогеогр. 2009 г.; 36 (8): 1573–1587. [Академия Google]
24. Д’Андреа Л., Мейрманс П., ван де Виль С., Гуаданьоло Р., ван Треурен Р., Козловски Г. и соавт.
Молекулярная биогеография салата колючего ( Lactuca serriola L.) показывает следы недавнего расширения ареала. Журнал наследственности. 2017; 108(2): 194–206. 10.1093/jhered/esw078
[PubMed] [CrossRef] [Google Scholar]
25. Министерство окружающей среды и энергетики. Влияние изменения климата в Victoria Online: Департамент окружающей среды и энергетики;
2017. Доступно по адресу: http://www.environment.gov.au/climate-change/climate-science-data/climate-science/impacts/vic. По состоянию на 10 сентября 2017 г. [Google Scholar]
По состоянию на 10 сентября 2017 г. [Google Scholar]
26. Чаухан Б.С., Джонсон Д.Е. Рост и размножение джунгрисиса ( Echinochloa colona ) в ответ на водный стресс. Сорняк наук. 2010 г.; 58(2): 132–135. [Google Scholar]
27. Каур С., Аулах Дж., Джала А.Дж. Рост и производство семян устойчивой к глифосату гигантской амброзии ( Ambrosia trifida L.) в ответ на водный стресс. Может J Plant Sci. 2016; 96(5): 828–836. [Google Scholar]
28. Bajwa AA, Chauhan BS, Adkins S. Морфологические, физиологические и биохимические ответы двух австралийских биотипов Parthenium hysterophorus к различным режимам влажности почвы. Environ Sci Pollut Res Int. 2017; 24 (19): 16186–16194. 10.1007/с11356-017-9176-1
[PubMed] [CrossRef] [Google Scholar]
29. Дюбуа М., Жиль К.А., Гамильтон Дж.К., Реберс П.Т., Смит Ф. Колориметрический метод определения сахаров и родственных им веществ. Анальная хим. 1956 год; 28(3): 350–356. [Google Scholar]
30. Lee S-S, Kim J-H. Общее количество сахаров, активность α-амилазы и прорастание после праймирования нормальных и состаренных семян риса. Корейский журнал растениеводства. 2000 г.; 45(2): 108–111. [Академия Google]
Lee S-S, Kim J-H. Общее количество сахаров, активность α-амилазы и прорастание после праймирования нормальных и состаренных семян риса. Корейский журнал растениеводства. 2000 г.; 45(2): 108–111. [Академия Google]
31. Джаванмарди Дж., Стушнофф С., Локк Э., Виванко Дж.М. Антиоксидантная активность и общее содержание фенолов иранских образцов Ocimum . Пищевая хим. 2003 г.; 83(4): 547–550. [Google Scholar]
32. Team R Core. R: Язык и среда для статистических вычислений. Вена, Австрия: R Foundation for Statistical Computing; 2016. Доступно по адресу: https://www.r-project.org/about.html. По состоянию на 10 марта 2018 г. [Google Scholar]
33. Manivannan P, Jaleel CA, Sankar B, Kishorekumar A, Somasundaram R, Lakshmanan GMA, et al.
Рост, биохимические модификации и метаболизм пролина в Helianthus annuus L., вызванный стрессом от засухи. Коллоиды Surf B Биоинтерфейсы. 2007 г.; 59(2): 141–149. 10.1016/j.colsurfb.2007.05.002
[PubMed] [CrossRef] [Google Scholar]
34. Чаухан Б.С. Реакция роста Itchgrass ( Rottboellia cochinchinensis ) на водный стресс. Сорняк наук. 2017; 61(1): 98–103. [Google Scholar]
Чаухан Б.С. Реакция роста Itchgrass ( Rottboellia cochinchinensis ) на водный стресс. Сорняк наук. 2017; 61(1): 98–103. [Google Scholar]
35. Кусака М., Лалусин А.Г., Фуджимура Т. Поддержание роста и тургора сортов проса африканского ( Pennisetum glaucum [L.] Leeke) с различной корневой структурой и осморегуляцией в условиях засушливого стресса. Наука о растениях. 2005 г.; 168(1): 1–14. [Академия Google]
36. Бхатт Р., Рао Н.С. Влияние загрузки бобов на реакцию бамии на водный стресс. Индийский J Plant Physiol. 2005 г.; 10(1): 54. [Google Scholar]
37. Саранги Д., Ирмак С., Линдквист Дж.Л., Кнежевич С.З., Джала А.Дж. Влияние водного стресса на рост и плодовитость водяной конопли ( Amaranthus rudis ). Сорняк наук. 2016; 64(1): 42–52. [Google Scholar]
38. Карими М., Ахмади А., Хашеми Дж., Аббаси А., Таварини С., Гульельминетти Л. и др.
Влияние истощения почвенной влаги на стевию ( Stevia rebaudiana Bertoni ), выращенных в тепличных условиях: рост, содержание стевиоловых гликозидов, растворимых сахаров и общая антиоксидантная способность. Научный Хортик. 2015 г.; 183:93–99. [Google Scholar]
Научный Хортик. 2015 г.; 183:93–99. [Google Scholar]
39. Li S., Pezeshki SR, Goodwin S. Влияние режимов влажности почвы на фотосинтез и рост рогоза ( Typha latifolia ). Акта Эколо. 2004 г.; 25(1): 17–22. [Google Scholar]
40. Мортенсен Д.А., Кобл Х.Д. Влияние влажности почвы на дурнишник обыкновенный ( Xanthium strumarium ) вмешательство в соевые бобы ( Glycine max ). Сорняк наук. 1989 год; 37(1): 76–83. [Google Scholar]
41. Webster TM, Gray TL. Рост и размножение бенгальского дневного цветка ( Commelina benghalensis ) в ответ на стресс от засухи. Сорняк наук. 2008 г.; 56(4): 561–566. [Google Scholar]
42. Пассиура Дж. Роль характеристик корневой системы в засухоустойчивости сельскохозяйственных культур. Доклад, представленный на Специальном международном симпозиуме по принципам и методам повышения устойчивости сельскохозяйственных культур к засухе: с акцентом на рис, Колледж, Лагуна (Филиппины), 4–8 мая 19 г.81.
43. Джалил К.А., Гопи Р., Санкар Б., Гоматинаягам М., Паннирселвам Р. Дифференциальная реакция эффективности использования воды у двух разновидностей Catharanthus roseus в условиях засушливого стресса. C R Биол. 2008 г.; 331(1): 42–47. 10.1016/j.crvi.2007.11.003
Джалил К.А., Гопи Р., Санкар Б., Гоматинаягам М., Паннирселвам Р. Дифференциальная реакция эффективности использования воды у двух разновидностей Catharanthus roseus в условиях засушливого стресса. C R Биол. 2008 г.; 331(1): 42–47. 10.1016/j.crvi.2007.11.003
[PubMed] [CrossRef] [Google Scholar]
44. Kramer PJ, Boyer JS. Водные отношения растений и почв. Нью-Йорк: Академическая пресса; 1995. [Google Scholar]
45. Yin C, Wang X, Duan B, Luo J, Li C. Ранний рост, распределение сухого вещества и эффективность использования воды двух симпатриков populus видов, пострадавших от водного стресса. Бот Environment Exp. 2005 г.; 53(3): 315–322. [Google Scholar]
46. Zhang M, Duan L, Zhai Z, Li J, Tian X, Wang B, et al. Влияние регуляторов роста растений на потери урожая сои, вызванные дефицитом воды. Материалы 4-го международного конгресса по растениеводству, Брисбен, Австралия; 2004.
47. Паттерсон Д.Т., Хайсмит М.Т. Соревнование шпорцевой аноды ( Anoda cristata ) и Бархатного листа ( Abutilon theophrasti 9)0030) с хлопком ( Gossypium hirsutum ) во время имитации засухи и восстановления. Сорняк наук. 2017; 37(5): 658–664. [Google Scholar]
Сорняк наук. 2017; 37(5): 658–664. [Google Scholar]
48. Chaves MM, Flexas J, Pinheiro C. Фотосинтез в условиях засухи и солевого стресса: механизмы регуляции от всего растения к клетке. Энн Бот. 2009 г.; 103(4): 551–560. 10.1093/аоб/mcn125
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
49. Flexas J, Bota J, Galmés J, Medrano H, Ribas-Carbó M. Поддержание положительного баланса углерода в неблагоприятных условиях: реакция фотосинтеза и дыхания к водному стрессу. Завод Физиол. 2006 г.; 127(3): 343–352. [Академия Google]
50. Fischer C, Höll W. Пищевые запасы сосны обыкновенной ( Pinus sylvestris L.). Деревья. 1991 год; 5(4): 187–195. [Google Scholar]
51. Мохаммадхани Н., Хейдари Р. Вызванное засухой накопление растворимых сахаров и пролина в двух сортах кукурузы. World Appl Sci J. 2008; 3(3): 448–453. [Google Scholar]
52. Al Hakimi A, Monneveux P, Galiba G. Растворимые сахара, пролин и относительное содержание воды (RCW) как признаки улучшения засухоустойчивости и дивергентный отбор для RCW от Triticum polonicum в Triticum durum .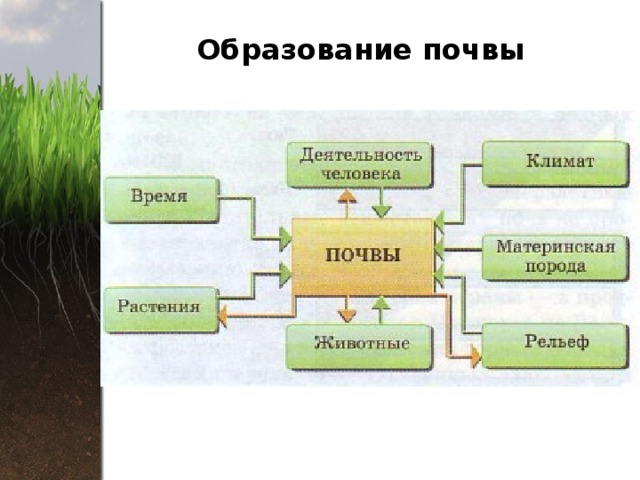 J Genet Breed (Италия). 1995. [Google Scholar]
J Genet Breed (Италия). 1995. [Google Scholar]
53. Хукстра Ф.А., Головина Е.А., Буйтинк Дж. Механизмы устойчивости растений к высыханию. Тенденции Растениевод. 2001 г.; 6(9): 431–438. [PubMed] [Google Scholar]
54. Камели А., Лозел Д. Углеводы и водный статус растений пшеницы в условиях водного стресса. Новый Фитол. 1993 год; 125(3): 609–14. [Google Scholar]
55. Осава Т. Новые природные антиоксиданты для использования в пищевых и биологических системах В: Uritani I, Garcia V, Mendoza E, editors. Послеуборочная биохимия растительных пищевых материалов в тропиках
Япония: Издательство Японских научных обществ; 1994. стр. 241–251. [Google Scholar]
56. Алексиева В., Сергиев И., Мапелли С., Каранов Е. Влияние засухи и ультрафиолетового излучения на маркеры роста и стресса у гороха и пшеницы. Окружающая среда растительной клетки. 2001 г.; 24 (12): 1337–1344. [Google Scholar]
57. D’Souza MR, Devaraj V. Специфические и неспецифические реакции гиацинтовых бобов ( Dolichos lablab ) на стресс от засухи. Индийская J Биотехнология. 2011 г.; 10(1): 130–139. [Google Scholar]
Индийская J Биотехнология. 2011 г.; 10(1): 130–139. [Google Scholar]
58. Уолк Дж.Л., Хидаяти С.Н., Диксон К.В., Томпсон К., Пошлод П. Изменение климата и регенерация растений из семян. Глоб Чанг Биол. 2011 г.; 17(6): 2145–2161. [Академия Google]
59. Норрис РФ. Echinochloa crus-galli (трава ежовника) семена дождя в условиях орошения. АСП БИОЛ. 2003 г.; 69: 163–170. [Google Scholar]
Изменение климата усилит жажду растений и нас
Совместное воздействие повышения уровня CO2 и повышения температуры увеличит потребление воды растениями. Это приведет к снижению уровня воды в ручьях и реках, таких как река Ашепу в Южной Каролине, изображенная здесь.
Фотография Винсента Дж. Муси, Коллекция изображений Nat Geo
Пожалуйста, соблюдайте авторские права. Несанкционированное использование запрещено.
К концу века растения смогут потреблять значительно больше воды, оставляя меньше воды людям в Северной Америке, Европе и Центральной Азии, даже если будет больше дождя и снега, сообщается сегодня в новом исследовании, опубликованном в журнале Nature Geoscience .
Растения являются основными регуляторами круговорота воды, ответственными за 60 процентов потока воды из земли в атмосферу. Исследования показывают, как изменение климата меняет этот жизненный цикл несколькими различными способами.
«Растения подобны атмосферной соломе, доминирующей в том, как вода течет из земли в атмосферу», — говорит географ-климатолог Джастин Манкин из Дартмутского колледжа и ведущий автор исследования.
Без значительного сокращения выбросов двуокиси углерода в ближайшие десятилетия средняя глобальная температура повысится на 4–6 градусов Цельсия, а к концу столетия содержание CO 2 в атмосфере почти удвоится. Те более горячие, богатые CO 2 будущие условия сродни включению тепла и перекачиванию CO 2 в теплицу. Вероятным результатом, при условии отсутствия других ограничивающих факторов, таких как нехватка питательных веществ, является взрыв растительной жизни. Но это оставит значительно меньше воды для использования людьми, сказал Манкин в интервью.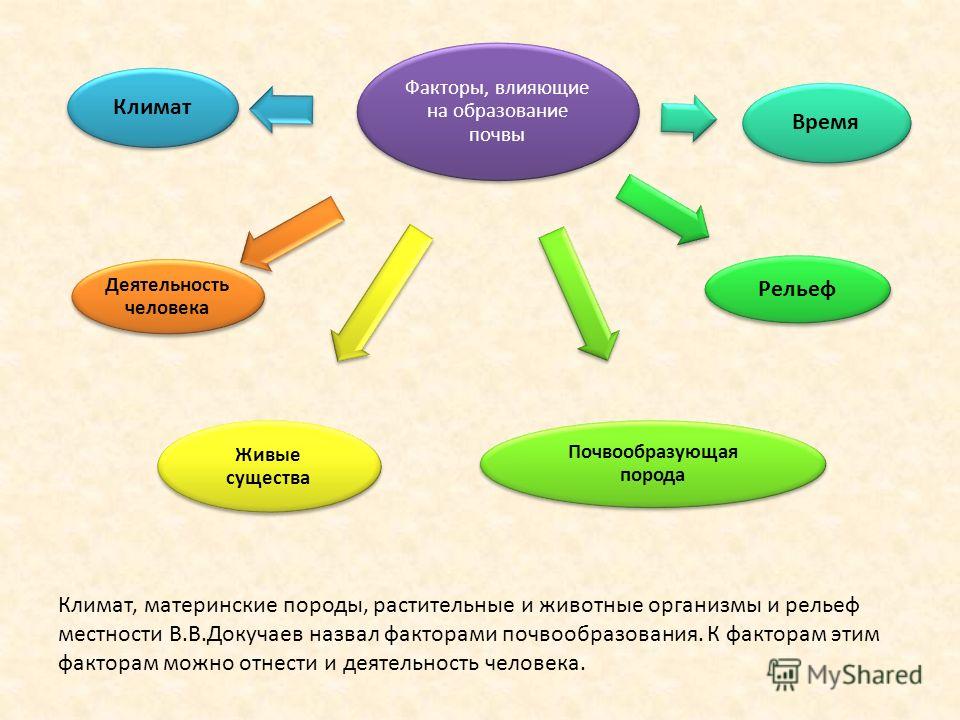 ( Прочтите об усилиях по защите мировой воды. )
( Прочтите об усилиях по защите мировой воды. )
Изменение климата влияет на рост растений тремя способами. Во-первых, по мере увеличения уровня CO 2 растениям требуется меньше воды для фотосинтеза. Долгое время считалось, что этот хорошо задокументированный эффект означает, что в почвах и ручьях будет больше пресной воды. Но второй эффект противоречит этому: потепление мира означает более длинные и теплые вегетационные периоды, что дает растениям больше времени для роста и потребления воды, высушивая землю.
Исследователи обнаружили третий эффект: повышение уровня CO 2 усиливает фотосинтез. Растения в этой более жаркой, богатой CO 2 среде вырастают крупнее, с большим количеством листьев. Это означает, что во время дождя будет гораздо больше мокрых листьев, что создаст большую площадь поверхности для большего испарения. По словам Манкина, компьютерное моделирование показывает, что такое усиленное испарение листьев оказывает большое влияние на сток и влажность почвы.
Команда Манкина использовала 16 различных климатических моделей с историческими данными для ряда переменных, включая осадки, испарение листьев, испарение почвы, индекс площади листьев, влажность почвы и т. д., которые точно воспроизводят прошлые условия. Будущие климатические переменные, такие как температура приземного воздуха и CO 2 уровня были добавлены, чтобы выяснить, как они повлияют на глобальный круговорот воды.
В то время как растения повсюду будут потреблять больше воды в более жарком, богатом CO 2 мире, в северных и тропических регионах прогнозируется достаточное количество осадков, чтобы компенсировать дополнительный рост растений, говорит Манкин.
Основной вывод исследования: совокупное воздействие повышенного содержания CO 2 и более высоких температур приведет к увеличению потребления воды растительностью. Это приведет к снижению уровня воды в реках и ручьях средних широт, включая Северную Америку, Европу и Среднюю Азию.
Плохие новости о воде
Уже давно ведутся споры о том, означает ли воздействие высокого уровня CO 2 на растения увеличение количества воды на земле, говорит Питер Глейк, всемирно известный эксперт по водным ресурсам и бывший президент Тихоокеанского института. который работает над глобальными водными проблемами.
«Благодаря более точному моделированию роста биомассы в целом, включая лиственный полог», исследование приходит к «надежному, противоположному и «плохому» выводу: повышение уровня CO 2 и связанные с этим изменения климата ухудшат, а не улучшат доступность воды», — говорит Глейк, не участвовавший в исследовании.
Этот результат «почти наверняка плохая новость для западной части США», — говорит он.
Предыдущее исследование климата выявило 80-процентную вероятность того, что к 2100 году юго-запад и центральная часть Великих равнин поразит 35-летнюю или более «мегазасуху» с обычными выбросами CO 2 . Умеренное сокращение выбросов снизит этот риск только до 60 процентов. И эта модель мегазасухи не включает новые данные о том, как изменения в растительности могут ухудшить условия, говорит Глейк.
И эта модель мегазасухи не включает новые данные о том, как изменения в растительности могут ухудшить условия, говорит Глейк.
Атмосфера уже стала более насыщенной CO 2 и климат стал теплее. По словам Манкина, спутниковые данные свидетельствуют о значительном увеличении растительности за последние 40 лет. В то время как вегетационные периоды также становятся длиннее, трудно сказать, что это недавнее озеленение Земли полностью связано с изменением климата, потому что за последние 100 лет ландшафт сильно изменился человеком, говорит он.
CO
2 уровни все выше и выше
В течение не менее 800 000 лет концентрация CO 2 в атмосфере колебалась от 180 до 290 частей на миллион (млн). За последние 10 000 лет они оставались на уровне 280 частей на миллион, пока промышленная революция не привела к широкому использованию угля.
Сегодняшние измерения показывают, что уровни CO 2 составляли 412 частей на миллион по состоянию на сентябрь этого года, что на 47 процентов выше доиндустриальных уровней.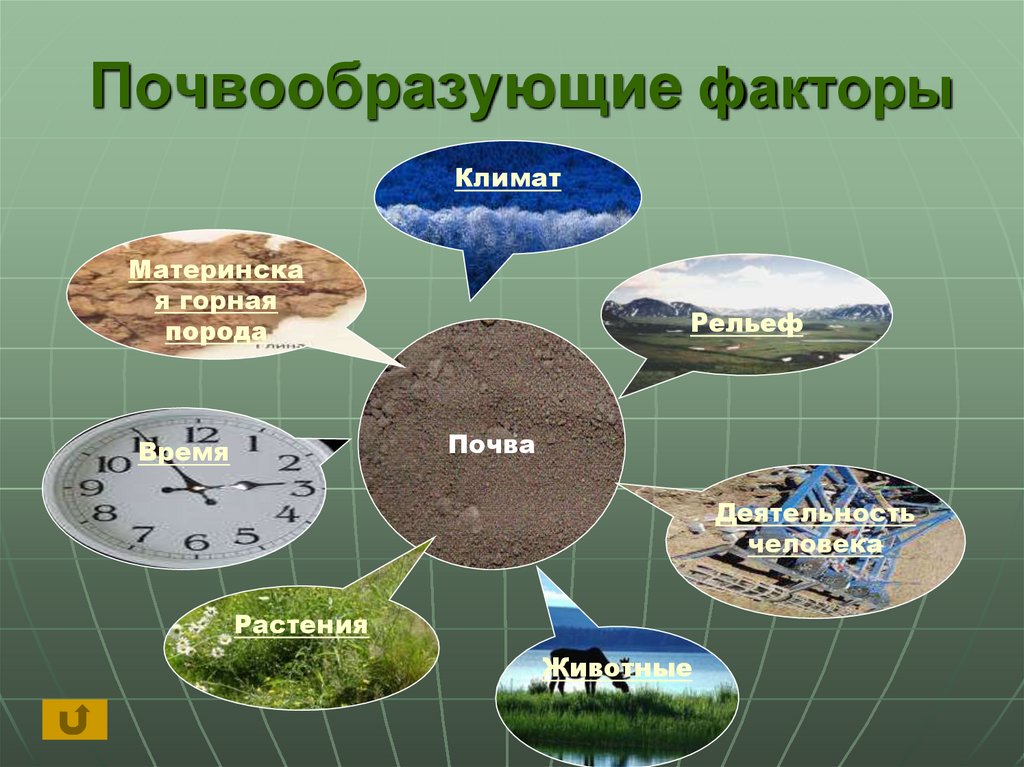 В последний раз уровни CO 2 превышали 400 частей на миллион от 16 до 25 миллионов лет назад, когда планета и ее климат были совсем другими.
В последний раз уровни CO 2 превышали 400 частей на миллион от 16 до 25 миллионов лет назад, когда планета и ее климат были совсем другими.
CO 2 уровни увеличиваются со скоростью 2 ppm в год. При продолжении использования угля, газа и нефти этот показатель может удвоиться до 560 частей на миллион к 2100 году. В этих условиях моделирование показывает, что засухи происходят намного быстрее, длятся дольше и становятся более сильными в средних широтах — даже при нормальных осадках. , — говорит Манкин.
Нехватка воды уже является серьезной проблемой: четыре миллиарда человек страдают от острой нехватки воды по крайней мере один месяц в году, согласно исследованию 2016 года. Любое сокращение подачи воды в будущем – это действительно очень плохие новости. Это верно даже для такой богатой страны, как США, говорит он, где уже есть люди от Детройта до юго-запада, страдающие от нехватки воды.
Читать дальше
Почему этот человек создает доспехи для кошек и мышей?
- История и культура
Зачем этот человек создает доспехи для кошек и мышей?
Художник Джефф де Бур десятилетиями проектировал и изготавливал крошечные доспехи музейного качества.