
Содержание
биофак СПбГУ
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Главная / Образование
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»>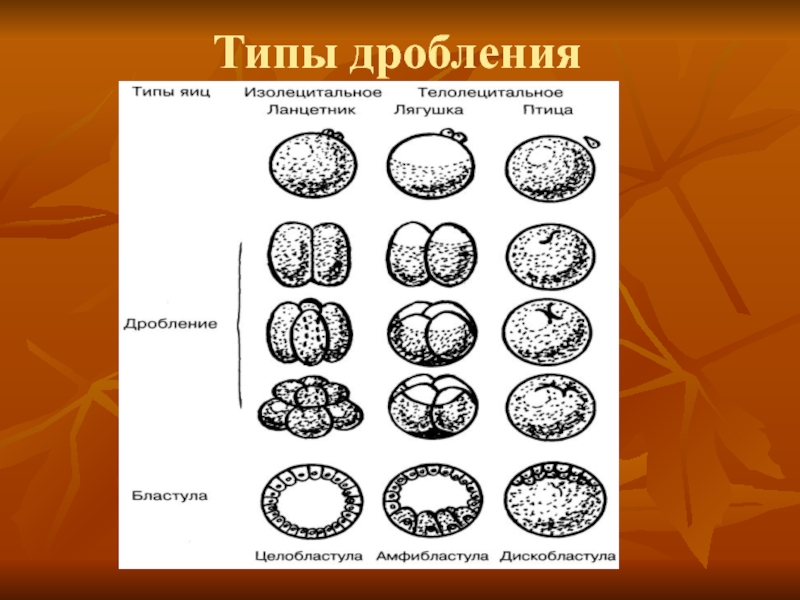 gif»>
gif»> gif»>
gif»> gif»>
gif»>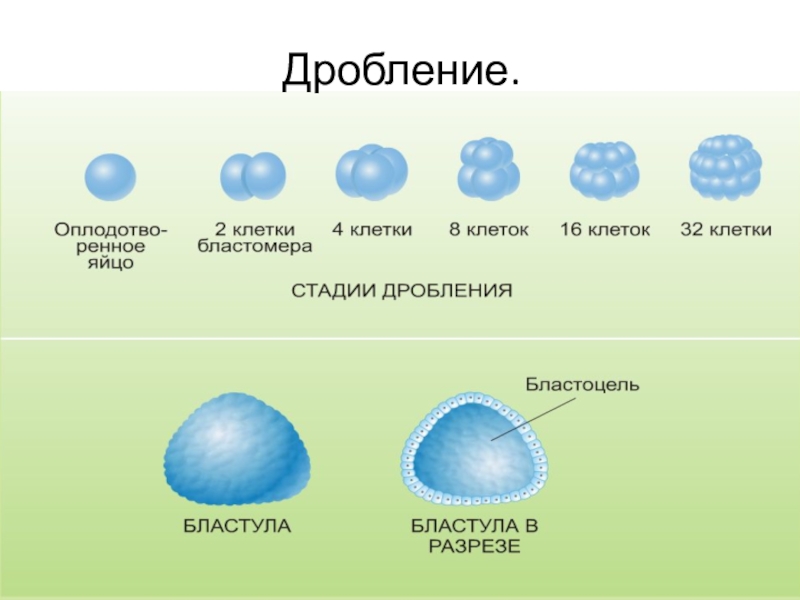 gif»>
gif»> gif»>
gif»> gif»>
gif»>контакты
карта сайта почтовый сервер
управление поддержка
199034, Санкт-Петербург, Университетская наб., 7-9
© Санкт-Петербургский государственный университет, 2006-2017
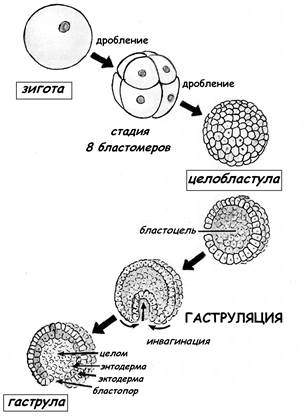
Типы дробления яйца
У разных видов животных дробление имеет свою особенную организацию, характер которой определяется прежде всего строением яйца, в частности распределением и количеством желтка, а также положением митотического веретена в бластомерах относительно анимально-вегетативной оси яйца.
Дробление бывает полным — голобластическим, неполным — меробластическим и поверхностным — абластическим. Полным называется дробление, при котором яйцо делится на бластомеры целиком. При неполном дроблении деления затрагивают только часть яйца, тогда как другая часть, обычно с высокой концентрацией желтка, не делится. При поверхностном типе дробления яйцеклетка не делится вовсе, и дело ограничивается лишь кариокинезом — делениями ядер.
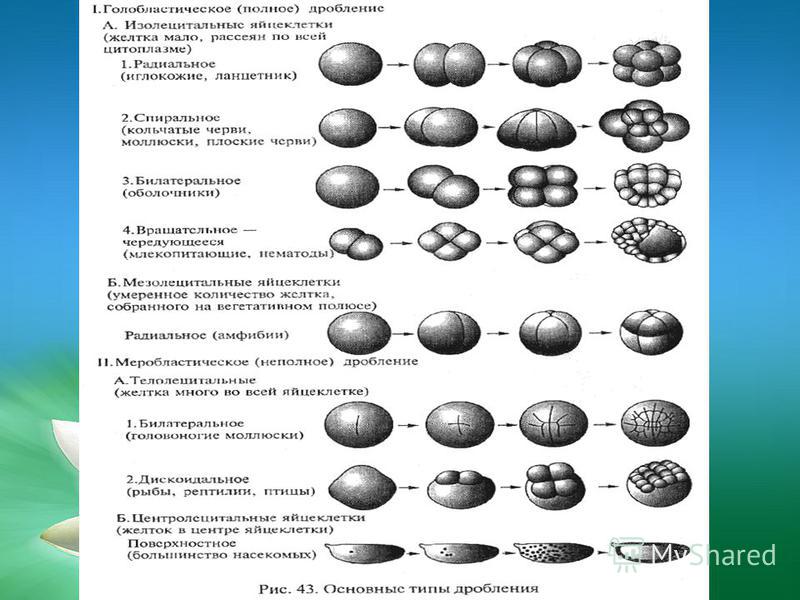
Голобластическое дробление. Такое дробление характерно для яиц, содержащих сравнительно мало желтка, т. е. для олиго- и мезолецитальных, а также для умеренно телолецитальных яиц. Если в результате дробления образуются бластомеры примерно одинаковых размеров, говорят о равномерном дроблении, если же бластомеры явно различаются по величине — о неравномерном. Неравномерность дробления может быть связана с концентрацией желтка в вегетативном полушарии. Иногда она обусловлена сосредоточением в отдельных бластомерах больших объемов специализированной цитоплазмы, например цитоплазмы полярной лопасти у некоторых моллюсков, или иными причинами, как в случае образования микромеров у морского ежа.
Меробластическое дробление. Этот тип дробления наблюдается у животных с телолецитальными яйцами, которые отличаются высокой степенью концентрации желтка в вегетативной области. У головоногих моллюсков, многих рыб, а также у рептилий и птиц дробление происходит только в относительно небольшой части яйца, образующей как бы диск на поверхности яйцеклетки, — дискоидальное дробление.
Абластическое дробление. Такое дробление характерно для центролецитальных яиц насекомых, поэтому его называют также централецитальным. В этом случае не происходит цитокинеза и деления цитоплазмы. Делятся только ядра, которые находятся в центральной области яйца, откуда они мигрируют по цитоплазматическим тяжам, пронизывающим яйцо, на поверхность. Длительное время зародыш имеет синцитиальную структуру. Попав в поверхностную цитоплазматическую бластему, или периплазму ядра образуют синцитиальную бластодерму, которая позднее целлюляризуется и дает начало клеточной бластодерме зародыша.
По характеру расположения бластомеров в развивающемся зародыше различают несколько типов дробления, в частности радиальное, бирадиальное, спиральное, билатеральное, ротационное и неупорядоченное дробление, а также табличную палинтомию и полиаксиальный тип дробления.
Радиальное дробление. У многих животных (книдарии, иглокожие, некоторые первично-хордовые, рыбы и амфибии) дробящееся яйцо имеет радиальную ось симметрии, при которой плоскость, проходящая через любой меридиан, делит зародыш на две геометрически тождественные половины. При радиальном дроблении два первых деления проходят во взаимно перпендикулярных меридиональных плоскостях, а третье — в экваториальной плоскости. Последующие деления чередуются в широтной и меридиональной плоскостях. Если третье деление происходит в экваториальной плоскости, то дробление равномерное, если же плоскость этого деления смещена в анимальное полушарие, то дробление неравномерное и ведет к образованию микромеров в анимальном и макромеров в вегетативном полушариях.
При радиальном дроблении два первых деления проходят во взаимно перпендикулярных меридиональных плоскостях, а третье — в экваториальной плоскости. Последующие деления чередуются в широтной и меридиональной плоскостях. Если третье деление происходит в экваториальной плоскости, то дробление равномерное, если же плоскость этого деления смещена в анимальное полушарие, то дробление неравномерное и ведет к образованию микромеров в анимальном и макромеров в вегетативном полушариях.
Спиральное дробление. У аннелид и моллюсков в результате первых двух взаимно перпендикулярных меридиональных делений образуется стадия четырех бластомеров. Начиная с третьего деления дробления митотические веретена располагаются под некоторым углом к меридиональной плоскости. Благодаря этому образующиеся четыре клетки анимального полушария несколько смешаются относительно клеток вегетативного квартета и располагаются в промежутках между его бластомерами квадрантами. Если смещение происходит по часовой стрелке (при наблюдении с анимального полюса), дробление называют дексиотропным (от лат. dexter — правый), если же смещение происходит в противоположном направлении, дробление называют леотропным. При последующих делениях наклоны веретен чередуются: за дексиотропным следует леотропное деление и наоборот. В случае спирального дробления его неравномерность может обнаружиться уже после первого деления дробления. Если на стадии 4 бластомеров все клетки одинаковых размеров, говорят о гомоквадрантном дроблении, если же они различаются по размерам, то — о гетероквадрантном.
dexter — правый), если же смещение происходит в противоположном направлении, дробление называют леотропным. При последующих делениях наклоны веретен чередуются: за дексиотропным следует леотропное деление и наоборот. В случае спирального дробления его неравномерность может обнаружиться уже после первого деления дробления. Если на стадии 4 бластомеров все клетки одинаковых размеров, говорят о гомоквадрантном дроблении, если же они различаются по размерам, то — о гетероквадрантном.
Неравномерностъ дробления может проявляться и вдоль анимально-вегетативной оси: при достаточно больших запасах желтка основной (вегетативный) квартет представлен макромерами, а анимальный — микромерами. В ходе спирального дробления бластомеры занимают строго фиксированное положение в системе и обозначаются специальными индексами.
Билатеральный тип дробления. Этот тип дробления характерен для нематод, а также для многих низших хордовых, в том числе для асцидий, аппендикулярий и бесчерепных. Характерной его особенностью является раннее проявление билатеральной симметрии.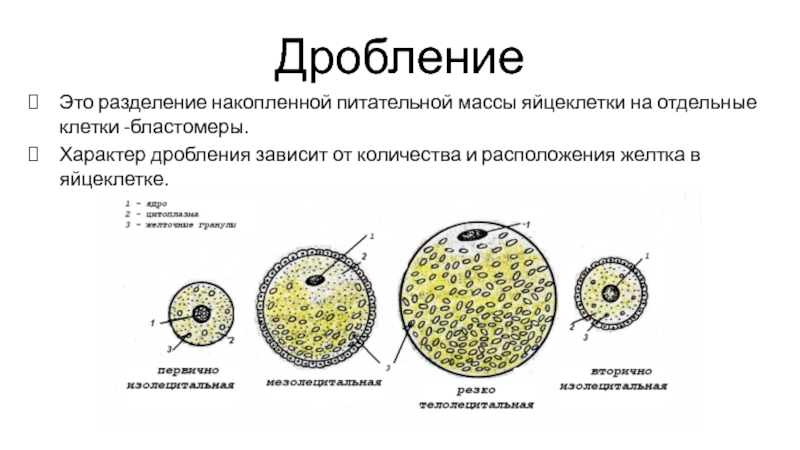 Например, у оболочников подразделение на левую и правую части происходит уже при первом меридиональном делении дробления, плоскость которого рассекает желтый серп оплодотворенного яйца на две симметричные половины. Билатеральность становится очевидной, когда вторая, тоже меридиональная борозда отделяет крупные передние бластомеры от задних более мелких клеток.
Например, у оболочников подразделение на левую и правую части происходит уже при первом меридиональном делении дробления, плоскость которого рассекает желтый серп оплодотворенного яйца на две симметричные половины. Билатеральность становится очевидной, когда вторая, тоже меридиональная борозда отделяет крупные передние бластомеры от задних более мелких клеток.
Некоторые авторы выделяют в особый, ротационный, тип дробления млекопитающих, у которых бластомеры при втором дроблении делятся во взаимно перпендикулярных плоскостях. Наконец, у некоторых животных описано вращение бластомеров. Изменение положения бластомеров относительно анимально-вегетативной оси, вероятно, характерно для ряда кишечнополостных. Согласно некоторым исследованиям, у книдарий иногда наблюдается слабая взаимосвязь между клетками, которая приводит к возникновению так называемого анархического, или неупорядоченного дробления.
Что определяет характер дробления? От чего зависит положение плоскости дробления? Что является причиной того или иного расположения митотического веретена? Конечно, характер дробления, как мы видели, отчасти определяется особенностями распределения желтка в яйцеклетке: увеличение концентрации желтка обычно приводит к замедлению образования борозд в вегетативном полушарии или даже к полному прекращению делений. В этих случаях наблюдается меробластическое или абластическое дробление.
В этих случаях наблюдается меробластическое или абластическое дробление.
Вместе с тем очевидно, что особенности того или иного типа дробления являются результатом длительной эволюции и контролируются не только концентрацией желтка, но многими факторами, природа которых во многом остается еще не раскрытой.
Сравнение разных типов дробления у животных позволило выдающемуся немецкому исследователю конца XIX — начала XX столетия О. Гертвигу вывести эмпирические правила («правила Гертвига»), которые, как первоначально казалось, дают ключ к пониманию причины того или иного положения плоскости деления яйца или бластомеров. Согласно одному из этих правил, ядро в яйцеклетке или бластомере занимает центр «активной» (то есть незагруженной желтком) цитоплазмы. Таким образом, при асимметричном расположении активной цитоплазмы предопределяется и аналогичное положение ядра. Согласно другому правилу Гертвига, ось веретена деления совпадает с направлением наибольшей протяженности цитоплазмы. Во многих случаях правила, установленные Гертвигом, оправдываются. Тем не менее они верны не всегда и, очевидно, будучи лишь результатом визуального анализа, не раскрывают реальных механизмов детерминации плоскостей дробления.
Во многих случаях правила, установленные Гертвигом, оправдываются. Тем не менее они верны не всегда и, очевидно, будучи лишь результатом визуального анализа, не раскрывают реальных механизмов детерминации плоскостей дробления.
Выполненные в конце 1930-х годов известным шведским эмбриологом Свеном Герстадиусом (1898-1996) опыты на морском еже показали, что положение веретен в последовательные моменты дробления предопределяется какими-то процессами в цитоплазме, которые активируются после оплодотворения. Герстадиус обнаружил, что, помещая яйца морского ежа в гипотоническую среду, или подвергая их встряхиванию, можно задержать то или иное деление дробления. При этом оказалось, что время появления того или иного положения веретена относительно момента оплодотворения остается неизменным. Например, если задержать первое деление на один цикл, то экваториальное деление, которое в норме является третьим, произойдет на стадии двух бластомеров, хотя при нормальном развитии для этой стадии характерно меридиональное деление. Как известно, микромеры вегетативного полушария у морского ежа возникают при четвертом делении дробления, т. е. на стадии образования 16-клеточного зародыша. Однако при задержке третьего деления дробления формирование микромеров происходит строго «по расписанию», т. е. через такое же время после оплодотворения, что и при нормальном развитии, но на стадии 4 бластомеров, а не 8, как при нормальном развитии.
Как известно, микромеры вегетативного полушария у морского ежа возникают при четвертом делении дробления, т. е. на стадии образования 16-клеточного зародыша. Однако при задержке третьего деления дробления формирование микромеров происходит строго «по расписанию», т. е. через такое же время после оплодотворения, что и при нормальном развитии, но на стадии 4 бластомеров, а не 8, как при нормальном развитии.
Из опытов Герстадиуса вытекало два важных следствия. Во-первых, они свидетельствовали, что в дробящемся яйце имеется некий счетчик времени, который, будучи независимым, тем не менее сопряжен с процессами, непосредственно контролирующими кариокинез. Во-вторых, эти опыты указывали на существование механизма, который предопределяет последовательный ряд событий, реализующих генетически детерминированную ориентацию веретен деления.
Важный вклад в понимание проблемы дробления примерно в то же время внесла американский эмбриолог Этель Гарвей (1885-1968), работы которой также показали, что механизмы кариокинеза и цитокинеза относительно автономны и могут быть разобщены в эксперименте. Центрифугируя неоплодотворенные яйца морского ежа в растворе сахарозы той же плотности, что и яйца, она смогла разделить яйцо на две половины — мерогоны. В легкой половине оставалось ядро, а в тяжелой концентрировался желток и пигментные зерна. После активации гипертонической морской водой наблюдалась партеногенетическая мерогоиия: безъядерные половинки начинали дробиться и формировали аномальные бластулы, которые после вылупления погибали, поскольку составляющие их клетки не имели ядер.
Центрифугируя неоплодотворенные яйца морского ежа в растворе сахарозы той же плотности, что и яйца, она смогла разделить яйцо на две половины — мерогоны. В легкой половине оставалось ядро, а в тяжелой концентрировался желток и пигментные зерна. После активации гипертонической морской водой наблюдалась партеногенетическая мерогоиия: безъядерные половинки начинали дробиться и формировали аномальные бластулы, которые после вылупления погибали, поскольку составляющие их клетки не имели ядер.
Относительная автономия кариокинеза и цитокинеза обусловлена тем, что в их основе лежат различные клеточные механизмы. Поскольку эти механизмы и, в частности, роль цитоскелета в осуществлении клеточного деления подробно рассматриваются в курсе цитологии, здесь мы ограничимся лишь кратким упоминанием о них.
Кариокинез происходит при участии митотического веретена, основу которого составляют микротрубочки, построенные из белка тубулина. Цитокинез осуществляется сократительным кольцом, которое образуется в области борозды дробления из актиновых микрофиламентов. Если дробящееся яйцо обработать ингибитором сборки микротрубочек (например, колхицином или нокодазолом), то кариокинетические деления блокируются в метафазе. При действии ингибиторов формирования микрофиламентов (например, цитохалазина) деление ядер сохраняется, но подавляется цитокинез.
Если дробящееся яйцо обработать ингибитором сборки микротрубочек (например, колхицином или нокодазолом), то кариокинетические деления блокируются в метафазе. При действии ингибиторов формирования микрофиламентов (например, цитохалазина) деление ядер сохраняется, но подавляется цитокинез.
При нормальном развитии образование веретена и борозды дробления скоординированы. Уже давно была отмечена связь между положением митотического веретена и направлением борозды дробления: последняя всегда проходит в плоскости метафазной пластинки, перпендикулярно длинной оси веретена. Экспериментальные воздействия, ведущие к изменению положения веретена, неизбежно вызывают смещение борозды дробления в соответствии с новым положением веретена. Неудивительно поэтому, что появление сверхчисленных веретен (например, при полиспермии) резко нарушает нормальный ход развития, а разрушение звезд веретена ведет к остановке дробления. Возможно, однако, что образование борозды дробления зависит главным образом от взаимодействия двух звезд, поскольку в экспериментальных условиях удается вызвать появление борозды между звездами, не связанными между собой веретеном.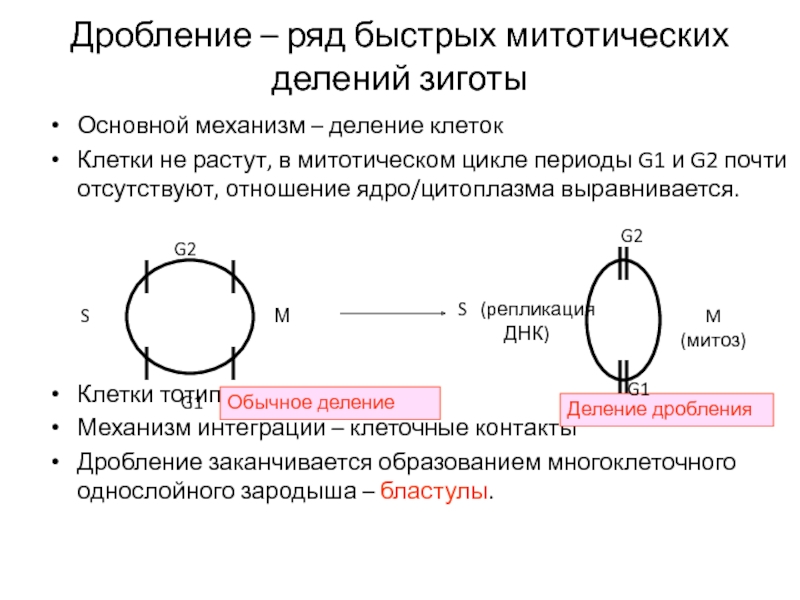 Можно предполагать, что область формирования сократительного кольца микрофиламентов определяется взаимодействием микротрубочек звезд с кортексом яйца.
Можно предполагать, что область формирования сократительного кольца микрофиламентов определяется взаимодействием микротрубочек звезд с кортексом яйца.
Положение митотического веретена, а следовательно, и звезд зависит не только от элементов цитоскелета, но и от других цитоплазматических факторов, хотя реальный механизм их действия пока не известен. Например, у животных со спиральным типом дробления обнаружено, что ориентация плоскости третьего деления в правую или левую сторону (т. е. дексиотропность или леотропность) контролируется цитоплазматическим фактором, который синтезируется под контролем генов материнского организма в период оогенеза. Значение этого фактора было показано как в опытах, в которых цитоплазму из яиц одного типа инъецировали в яйца другого типа, так и в генетических исследованиях закрученности раковины у моллюска Lymnea, которая коррелирует с ориентацией веретена при дроблении. Так, у Lymnea аллель правозакрученности доминантен (D), а левозакрученности рецессивен (d). При скрещивании самок DD с самцами dd все потомки Dd были правозакрученными. При скрещивании самцов DD с самками dd все потомки Dd были левозакрученными, а при скрещивании самцов Dd с самками Dd потомки (1DD: 2Dd: 1dd) были все правозакрученными.
При скрещивании самок DD с самцами dd все потомки Dd были правозакрученными. При скрещивании самцов DD с самками dd все потомки Dd были левозакрученными, а при скрещивании самцов Dd с самками Dd потомки (1DD: 2Dd: 1dd) были все правозакрученными.
Прямые доказательства участия генетических факторов в определении характера дробления получены на нематоде Caenorhabditis elegans, у которой первое деление дробления дает крупный передний бластомер АВ и задний бластомер Р1 меньших размеров. При втором делении образуется характерная Т-образная фигура, так как оси митотических веретен в указанных бластомерах располагаются взаимно перпендикулярно. Асимметрия деления яйца у С, elegans определяется белками PAR и МЕХ, локализованными в кортикальном слое. Обнаружено 6 генов par (partitioning-defective). У большинства мутантов par первое деление равномерное, а веретена дочерних клеток расположены вдоль одной оси, а не взаимно перпендикулярны. Эти нарушения коррелируют с изменениями локализации белков PAR. У животных дикого типа белки PAR-3 и PAR-6 локализованы в передней области кортекса, тогда как PAR-1 и PAR-2 — в задней. Отсутствие одного из белков передней группы ведет в распространению задних факторов вперед. В отсутствии PAR-2 факторы переднего комплекса распространяются назад.
У животных дикого типа белки PAR-3 и PAR-6 локализованы в передней области кортекса, тогда как PAR-1 и PAR-2 — в задней. Отсутствие одного из белков передней группы ведет в распространению задних факторов вперед. В отсутствии PAR-2 факторы переднего комплекса распространяются назад.
В настоящее время эмбриология не располагает данными, которые позволили бы дать сколько-нибудь связный очерк механизмов, лежащих в основе формирования тех или иных конкретных типов дробления. Тем не менее даже те отрывочные сведения, которые имеются в нашем распоряжении, позволяют утверждать, что тип дробления определяется сложным комплексом факторов, в том числе генетическими и клеточными механизмами, которые контролируют и интегрируют многие элементарные процессы. Среди последних особое значение имеют события, связанные с синтезом ДНК, организацией митотического деления ядра (кариокинез) и делением цитоплазматического тела клетки (цитокинез). Относительная автономия этих элементарных процессов создает предпосылки для формирования в ходе эволюции животных разнообразных типов дробления. Например, в основе возникновения поверхностного типа дробления лежит возможность разобщения кариокинеза и цитокинеза.
Например, в основе возникновения поверхностного типа дробления лежит возможность разобщения кариокинеза и цитокинеза.
Роторный дробильный клапан
— Hanningfield
Техническое описание
Роторная дробилка, изготовленная в соответствии со стандартами cGMP, способна дробить продукты одним или двумя роликами на этапе обработки.
Этот инструмент подходит для систем безразборной мойки и стерилизации на месте, имеет компактную стандартную конструкцию и может функционировать как предварительный измельчитель в сочетании с конусной мельницей или другой машиной ситового типа. .
Материалы
> Нержавеющая сталь EN 1.4404 (AISI 316L)
Motion Drive
> Электродвигатель
Отделка
> Ra 0 0,5 мкм внутренний
> Ra 1,2 мкм внешний
Декларации и сертификаты
> Сертификация FDA (CFR § 177.2600)
> Сертификат материала 3. 1 согласно EN10204
1 согласно EN10204
Размеры
DN 100, DN 150, DN 200, DN 250, DN 300
Загрузки
Выбрать другую модель
Поворотные затворы
являются одними из наиболее распространенных клапанов, используемых в технологических процессах.
Гибкий клапан по своей сути такой же, как дроссельный клапан, но вместо лопасти из нержавеющей стали гибкий клапан оснащен силиконовой лопастью.
Sterisplit — это раздельный дроссельный клапан или клапан с высокой степенью регулирования, предназначенный для безопасного перемещения порошков в стерильных или потенциально опасных условиях.
Легкий поворотный клапан подходит для фармацевтической, химической и пищевой промышленности.
Rotovalve Plus предназначен для подачи и объемного дозирования сыпучих продуктов.
Поворотный клапан для тяжелых условий эксплуатации подходит для фармацевтической, химической и пищевой промышленности.
Rotodoser предназначен для применений, где требуется высокая точность и точное дозирование.
Мы понимаем, что стандартные клапаны, имеющиеся в наличии, не всегда подходят для вашего применения, поэтому мы предлагаем индивидуальные решения для наших клиентов.
Мы можем поставить все запасные и быстроизнашивающиеся детали, относящиеся к ассортименту Sterivalves, включая техническую документацию.
Запросить дополнительную информацию
Роторные дробилки
от Sturtevant Inc. Роторные дробилки
от Sturtevant Inc.
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Кубики размера корма
Кубики размера корма 