
Содержание
Генезис, характеристика, классификация почв. Материалы почвенных исследований и их использование
Арктическая и тундровая зоны входят в Евроазиатскую полярную область.
Условия почвообразования и почвы арктической зоны.Арктические почвы формируются в условиях полярного холодного сухого климата. Осадков выпадает 50-200 мм, температура июля не выше +5°С, безморозный период отсутствует. Почвы оттаивают на глубину 30-40 см. Среднегодовые температуры отрицательные – от -14 до -18°С. Растительность представлена мхами, лишайниками, различными видами водорослей. Высшие растения на водоразделах: занимают менее 25% территории. Количество опада – в пределах 0,5 т/га. Значительные площади занимает голый грунт. Большая роль в почвообразовании и формировании нанорельефа принадлежит криогенным процессам и физическому выветриванию.
Основным типом почв являются арктические, которые разделяют на два подтипа: арктические пустынные и арктические типичные гумусовые. Профиль состоит из двух горизонтов – А и С, иногда с переходным горизонтом АС. Для арктических почв характерны: небольшая мощность почвенного профиля, в пределах 30-40 см, скелетность, отсутствие оглеения, связанное с небольшим количеством осадков и просыханием почв под действием сильных ветров. Содержание гумуса в горизонте. А может достигать 2-4%, реакция среды – от слабокислой до слабощелочной, в зависимости от состава почвообразующих пород. Иногда почвы содержат карбонаты и водорастворимые соли.
Для арктических почв характерны: небольшая мощность почвенного профиля, в пределах 30-40 см, скелетность, отсутствие оглеения, связанное с небольшим количеством осадков и просыханием почв под действием сильных ветров. Содержание гумуса в горизонте. А может достигать 2-4%, реакция среды – от слабокислой до слабощелочной, в зависимости от состава почвообразующих пород. Иногда почвы содержат карбонаты и водорастворимые соли.
Условия почвообразования и почвы тундровой зоны. Климат тундровой зоны характеризуется холодной зимой, коротким летом. Осадков выпадает от 400 мм на Кольском полуострове до 150-250 мм в Восточной Сибири. В южной тундре средняя температура июля – 8-13°С. Относительная влажность воздуха достигает 80-90%. Средняя годовая температура колеблется от -2°С (на западе) до -14…-16С° (в азиатской части). Это зона вечной мерзлоты. Летом почва оттаивает на глубину от 30 см на болотных торфяных почвах и до 1-2 м на песчаных.
Растительность арктической тундры представлена злаково-осоково-моховыми ценозами, а в понижениях – гипново-осоковыми ассоциациями на полигональных болотах. Типичная тундра характеризуется господством мхов и лишайников. Мхи преобладают на суглинистых почвах, лишайники – на щебнистых.
Типичная тундра характеризуется господством мхов и лишайников. Мхи преобладают на суглинистых почвах, лишайники – на щебнистых.
При продвижении к югу, в лесотундру, начинают появляться кустарнички – карликовые березы, ива, вереск, багульник, голубика, а по долинам рек на песчаных и супесчаных почвах – изреженные, угнетенные леса (ель, береза, лиственница и др.).
Количество опада составляет 0,5-1 т/га. В составе опада – низкое содержание оснований и азота.
Почвообразующие породы представлены морскими, ледниковыми и аллювиальными отложениями. В формировании бугристого и пятнистого микро- и нанорельефа большая роль принадлежит криогенным процессам: трещинообразование, солифлюкция, термокарст и др.
Зональным типом почв являются тундровые глеевые почвы. Изучение генезиса тундрово-глеевых почв проводилось Ю.А. Ливеровским, Е.Н. Ивановой, И.В. Забоевой, Н.А. Караваевой, В.О. Таргульяном, В.Д. Василевской и другими учеными. Низкие температуры, короткий период биологической активности, поверхностное и внутрипочвенное (надмерзлотное) переувлажнение определяют направленность почвообразования в тундре. В формировании профиля тундровых глеевых почв принимают участие три группы процессов: гумусообразование (детритообразование), продуцирующее сухоторфянистый или грубогумусовый горизонт, оглеение и криогенез. Образование грубогумусовых и оторфованных горизонтов связано с пониженной биологической активностью и низким содержанием оснований и азота в составе опада.
В формировании профиля тундровых глеевых почв принимают участие три группы процессов: гумусообразование (детритообразование), продуцирующее сухоторфянистый или грубогумусовый горизонт, оглеение и криогенез. Образование грубогумусовых и оторфованных горизонтов связано с пониженной биологической активностью и низким содержанием оснований и азота в составе опада.
Тундровые глеевые почвы имеют следующее строение почвенного профиля: Ао – АоА1 – Вg –G. Мощность почвенного профиля небольшая, ограничивается мощностью деятельного (сезоннооттаивающего) слоя. Мощность органогенных горизонтов Ао+АоА1 может достигать 10-20 см. Содержание органического вещества в грубогумусовом горизонте составляет 5-10% и более, в Н оторфованном горизонте Ао – 30-60%. В составе гумуса преобладают фульвокислоты (СГК:СФК – 0,3-0,5). Отмечается повышенное содержание гумуса (1,5-2%) по всему профилю, связанное с потечностью гумуса и механической аккумуляцией над многолетнемерзлым водоупорным горизонтом. Реакция среды кислая и слабокислая, насыщенность основаниями – 20-50%.
Разделение тундрово-глеевых почв на подтипы и роды связано с характером органогенных горизонтов (грубогумусовый, торфянистый, перегнойный), оглеением (поверхностно-глеевые, надмерзлотно-глеевые), с начальными стадиями проявления элювиальных и иллювиальных ЭПП (оподзоленные, иллювиально-железисто-гумусовые).
Болотные почвы тундры представлены преимущественно низинными обедненными торфяными и торфяно-глеевыми, но встречаются и верховые.
Земледелие в тундре носит очаговый характер. Имеется опыт выращивания трав, капусты, моркови, лука, картофеля. Очаги земледелия приурочены к легким песчаным и супесчаным почвам, чаще всего – в долинах рек с глубоким деятельным слоем или отсутствием горизонтов вечной мерзлоты. Весьма перспективным для тундровой зоны является закрытый грунт – выращивание овощей в теплицах.
Комплексная географическая характеристика. Тундрово-глеевые почвы
1. Комплексная географическая характеристика. Тундрово-глеевые почвы.
Литвинов Я. В. ЭКП-1-2018-НМ
В. ЭКП-1-2018-НМ
2. Географический ареал
3. Распространение
Тундровые глеевые почвы типичны
для тундровой ландшафтногеографической зоны. Они тянутся
полосой различной ширины по всей
северной окраине Евразии и Северной
Америки. В южном полушарии из-за
отсутствия суши в соответствующих
широтах тундровые глеевые почвы не
распространены. В Евразии эти почвы
составляют 2,7% от площади
континента. В Северной Америке их
доля в почвенном покрове даже выше
— до 4,6%. Общая площадь их на
земном шаре около 2600 тыс. км2.
4. На почвенной карте мира
Краткая географическая
энциклопедия, Том 3/Гл.ред.
Григорьев А.А. М.:Советсвкая
энциклопедия — 1962, 580 с. с илл.,
19 л. карт
5. Характеристика условий почвообразования
6. Климат
Южная граница тундры приблизительно совпадает с июльской изотермой воздуха в 12°С. При средней июльской
температуре ниже 10—12° уже не могут расти деревья. Лета, в нашем понимании, если называть летними дни со
средней суточной температурой воздуха выше 12°, в тундре, как правило, не бывает. Самая теплая часть
Самая теплая часть
тундровой зоны расположена на Кольском полуострове. К востоку нарастает Континентальность климата тундры —
осадков становится меньше, а зимы холоднее. Мурманское побережье, находящееся под влиянием Гольфстрима,
имеет осадков 350—400 мм в год, средние температуры: февраль —6,2°, июль-август 9,1 °С, амплитуда — 15,3°,
тогда как в дельте реки Лены осадков всего 100 мм в год, средняя температура февраля —42°, а июля 5 °С, т. е.
амплитуда около 47°. За Колымой сказывается влияние Тихого океана, и климат снова приближается к
океаническому: зимы не так морозны, но лето прохладнее. Сумма активных температур по разным источникам
составляет 300 – 600 °С.
Основная часть тундрового сообщества деятельна лишь в летнее время (растительность, микроорганизмы,
беспозвоночные животные). Летом протекают и все основные абиотические процессы в ландшафте: выветривание,
эрозия, оттаивание мерзлоты и т. д. Поэтому первостепенное значение в жизни тундры имеет продолжительность
безморозного периода, который и определяет главные особенности тундрового ландшафта, его органического
мира. Этот период повсеместно менее 4 месяцев.
Этот период повсеместно менее 4 месяцев.
Общее количество осадков в тундре незначительное, в среднем 150—250 мм с отклонениями в меньшую и большую
стороны. По количеству осадков тундра приближается к пустынным регионам низких широт. Однако в тундре
много воды, высокая влажность почвы и воздуха. Большие территории заняты болотами. Тундра увлажнена
сильнее прочих ландшафтов Земли. Нигде ландшафтообразующая роль воды не выступает так отчетливо, как в
тундре. Подземные льды, снег, талые воды, туманы и длительные моросящие дожди — все это мощнейшие
экологические и ландшафтообразующие и почвообразующие факторы в тундре. Избыток воды связан с низкой
испаряемостью и транспирацией растениями, которая повсеместно не превышает 100 мм в год. Коэффициент
увлажнения в тундре всегда более 1,0.
7. Климатические пояса и области мира
Краткая характеристика: холодный,
сухой.
8. Границы субарктического климата по Алисову
Главной чертой субарктического
климата является если не
отсутствие, то низкая
продолжительность
климатического лета: даже в самый
тёплый месяц средняя температура
воздуха не превышает +15 °C.
Всегда возможны заморозки. Зима
всегда длительная, в зависимости
от местоположения может быть как
очень суровой, так и относительно
мягкой.
9. Тип растительности
Растительность тундры составляют в
первую очередь лишайники и мхи;
встречаются покрытосеменные
растения — невысокие травы
(особенно из семейства Злаки),
осоки, полярные маки и др.,
кустарники и кустарнички
(например, дриада, некоторые
карликовые виды берёзы и ивы,
ягодные кустарнички княженика,
голубика, морошка).
10. Тип растительности
В тундре выделяются три подзоны:
подзона южных кустарниковых (моховокустарниковых) тундр, подзона типичных
моховых (пушицево-моховых) тундр и
подзона арктических тундр. В отличие от
южных и типичных тундр для арктических
тундр характерна несомкнутость
растительного покрова; доминирующий
тип распределения растительности, как и
в арктической зоне, полигональносетчатый.
По химическому составу растительные
остатки отличаются исключительно низкой
зольностью. При их разложении
При их разложении
образуются органические кислоты,
вызывающие сильное подкисление
почвенной массы.
11. Материнская порода
Горные породы — ледниковые, морские и аллювиальные отложения различного
гранулометрического состава, часто сильно каменистые. В горах
почвообразующие породы представлены преимущественно грубоскелетным
элювием коренных пород.
Тип тундровых глеевых почв делится на два подтипа: 1) тундровые глеевые и 2)
тундровые глеевые оподзоленные. Тундровые глеевые почвы наиболее широко
развиты в полосе мохово-лишайниковых тундр и делятся на два рода:
тундровые глеевые (собственно) и тундровые глеевые иллювиально-гумусовые.
Тундровые глеевые (собственно) почвы формируются на суглинистых и
глинистых породах. Тундровы е глеевые иллювиально-гумусовые почвы
формируются на песчаных и супесчаных породах.
12. Основные почвообразовательные процессы
13. Развитие тундровых почв определяют главным образом процессы болотного почвообразования, протекающие в тундровых условиях
1. Оторфовывание органических остатков и их накопление на минеральной поверхности почвы.
Оторфовывание органических остатков и их накопление на минеральной поверхности почвы.
Торфообразование проявляется всегда, но в разной степени, от маломощных горизонтов до бугристых
торфянников, со слоем торфа около 50 см. Однако преобладающая мощность торфяного горизонта обычно
не превышает 20 см.
2. Гумусообразование с преимущественным образованием фульвокислот при разложении торфяного
горизонта, их миграция вместе с другими органическими соединениями по всему активному
надмерзлотному слою почвы при возможном накоплении в нижних горизонтах. Формирование гуминовых
темноокрашенных фракций гумуса крайне ограничено, а гумусовый горизонт фрагментарен по мощности.
Гумусовые вещества всегда кислые, ненасыщенные из-за крайне низкой зольности растительных остатков
тундровой растительности.
3. Оглеение всей минеральной толщи от торфяного слоя до вечной мерзлоты, с образованием мощного
глеевого горизонта G. Развитию анаэробного глеевого процесса способствуют передвижение минеральной
массы и постоянная высокая обеспеченность среды растворимыми органическими веществами из верхнего
оторфянного органогенного горизонта.
В относительно теплых местах тундры, преимущественно в долинах рек, возможно развитие настоящего
дернового процесса с образованием почв, близких к луговым. В южной тундре характерно появление
элементов подзолистого процесса с выделением особого подтипа почв лесотундровой подзоны.
14. Сущность процессов почвобразования
Оторфовывание – 1) накопление медленно гумифицирующихся и почти не
минерализующихся растительных остатков. Процесс протекает в анаэробной среде
при избыточном увлажнении; 2) консервация отмерших органических остатков при
незначительной гумификации.
Гумусообразование – разложение растительных остатков на месте их отмирания (in
situ) и последующего новообразования гумуса без его перемещения по профилю.
Оглеение. Термины «глей» и «глеевый процесс» ввел Г. Н. Высоцкий. Под глеем
понимали не очень плотную породу серого цвета с зеленоватым оттенком,
формирующуюся в условиях длительного переувлажнения. Глееобразование –
сложный биохимический восстановительный процесс (маслянокислое брожение),
протекающий в анаэробных условиях при обязательном присутствии органического
вещества и участии анаэробных микроорганизмов. Сущность процесса – под
Сущность процесса – под
воздействием неспецифических гетеротрофных анаэробных микроорганизмов Fe
свободных окисных соединений восстанавливается до закисного 2-х валентного,
вступает в комплексные связи с органическими соединениями и алюмосиликатами.
15. Строение профиля и процессы образования профилей. Основные черты каждого горизонта
16. Типичный профиль тундрово-глеевой почвы определяют следующие горизонты:
АО Cодержит слаборазложившиеся растительные остатки.
Соотношения органической и минеральной частей горизонта
варьируют. Органические остатки бурого и темно-бурого цвета,
разной степени измельчения и разложения. На поверхности
отдельных минеральных зерен — органо-железистые сгустки.
Встречаются гумусово-железистые рыхлые скопления (до 0,15–
0,20 мм), по периферии, по контуру покрытые оптически
ориентированными глинами. Микросложение очень рыхлое.
Вg Четко выражена микрозональность. Микрозоны оглеения
имеют компактное микросложение, светлую окраску плазменного
материала, чешуйчатое строение оптически
ориентированной плазмы. Микрозоны окисления характеризуются
Микрозоны окисления характеризуются
бурыми тонами, пленки и хлопья полуторных оксидов на зернах
скелета, пятна и микроконкреции железа. По границам микрозон
образуются тонкие трещины. Горизонт характеризуется
криогенной структурой или бесструктурный. Содержит пылеватый
материал, много глины. Органоглинистый материал находится как
в неагрегированном виде, так и в форме сгустков, комочков
округлой формы. Глины ориентируются по растительным остаткам
в форме полос.
G Плазма светло-серого цвета, материал не агрегирован.
Микросложение компактное с единичными порами, вокруг
которых формируются
слабоожелезненные кутаны ориентированной глины. Выражены
вертикальные зоны с признаками пропитки подвижным
органическим веществом. Плазма анизотропна, имеет чешуйчатое
или сетчатое строение. Возможно проявление криогенной
структуры
А0 — несколько оторфованная подстилка мощностью 3-5
см;
A1 — гумусовый (перегнойный или торфянистый)
горизонт мощностью 0-20 см, темно-серый или
коричнево-бурый, суглинистый, влажный,
переплетенный корнями растений, хорошо отслаивается
от других горизонтов; граница неровная, иногда этот
горизонт выклинивается;
Bg(G) — иллювиальный горизонт (или глеевый), иногда
подразделяется на подгоризонты, мощностью 40-55 см,
оглеенный, неравномерно окрашенный, на буром фоне
ржавые и сизые пятна, влажный, суглинистый, иногда
слоистый, часто тиксотропный; переход по границе
оттаивания;
GM — глеевый, мерзлый, темно-сизый, суглинистый, со
многими льдистыми прожилками.
18. Типичный профиль тундрово-глеевой почвы
Типичное строение тундровых почв как правило нарушается
мерзлотными явлениями криогенеза. Криогенез — комплекс
почвенных и грунтовых процессов, связанных с промерзанием и
оттаиванием. Присутствуют совокупность явлений физического,
химического и биологического преобразования почвенных масс.
Тиксотропия, солифлюкция, образование пятнистых и
полигональных тундр, гидролаколиты, термокарст и некоторые
другие явления — все это свойственно криогенезу. Тиксотропия —
явление превращения твердой почвенной массы в жидкотекучую
(геля в золь). Это происходит при механических воздействиях на
почву. С тиксотропией связана солифлюкция — сползание
тиксотропного почвенного слоя вниз по уклону под воздействием
силы тяжести. Глеевый почвенный слой разжижается и переходит в
плывунное состояние. Тиксотропность глеевого горизонта может
привести к формированию пятнистых тундр.
Специфический элемент тундровых ландшафтов — бугры-
гидролаколиты. Их высота меняется от 1 м (диаметром 2—5 м) до 70
Их высота меняется от 1 м (диаметром 2—5 м) до 70
м (диаметром 150—200 м). Возникновение бугров объясняют
пучением грунта в результате образования подземной ледяной
линзы. Оттаивание гидролаколитов по разным причинам, в
основном антропогенного происхождения, приводит к просадкам
почв и грунтов, которые называют термокарстом. При этом
образуются провалы, сдвиги, ямы, которые разрушают все
наземные сооружения, и в первую очередь дорожную сеть.
19. Физико-химические свойства
Количество гумуса, кислотность, уровень плодородия
Минеральный профиль тундровых глеевых почв практически
не дифференцирован по распределению ила и валовых
полуторных оксидов. Иногда проявляющееся обеднение
илом верхнего минерального горизонта может быть связано
с латеральным элювиированием. Характерно повышенное
содержание оксалаторастворимого железа в верхних
горизонтах. Реакция почв от кислой до слабокислой, емкость
поглощения невысокая, степень насыщенности основаниями
до 60% в верхнем минеральном горизонте и увеличивается
книзу. Наиболее кислыми являются органогенные
Наиболее кислыми являются органогенные
горизонты. Содержание гумуса в верхнем горизонте 4–6%,
характерна прогумусированность всего деятельного слоя,
иногда с образованием надмерзлотных максимумов. В
составе гумуса фульвокислоты преобладают над гуминовыми
кислотами (Сгк/Сфк 0,3–0,6). Тундровые глеевые почвы
характеризуются высокой плотностью, низкой порозностью,
низкой фильтрационной способностью глеевых горизонтов.
Усиливающиеся к югу процессы оглеения и нисходящей
миграции способствуют формированию в южных тундрах
тундровых глеевых оподзоленных почв. Последние несут
черты оподзоленности как в морфологии профиля, так и в
химическом составе. Они отличаются от тундровых глеевых
более кислой реакцией, меньшей насыщенностью
основаниями, элювиально-иллювиальной дифференциацией
минеральной толщи, более глубоким залеганием льдистой
многолетней мерзлоты (100–150 см).
Содержание гумуса до 5%. Крайне низкий уровень
плодородия.
21. Хозяйственное применение
Земледелие в тундровой зоне практически
невозможно. В ней распространено только
мелкое потребительское огородничество,
сеют репу, редьку, брюкву, лук и сажают
картофель. При этом решается проблемы
улучшения теплового режима, питания
растений, выбора «теплых» почв,
оптимизация обеспеченности растений
водой.
Территория тундры в
сельскохозяйственном отношении имеет
большое значение как кормовая база
северного оленеводства.
В этих зонах сосредоточено 41,6% всей
площади оленеводческих пастбищ страны.
Основные пастбища расположены в полосе
моховолишайниковых и кустарниковых
тундр. Лишайниковые тундры
используются как зимние пастбища, а
моховые, травяно-моховые и ерниковые
тундры — как летние. Арктическая тундра
менее благоприятна для оленеводства.
Сдвиги прокариотических сообществ при почвообразовании на песках в тундровой зоне
1. Esposito A, Ciccazzo S, Borruso L, Zerbe S, Daffonchio D, Brusetti L. Трехуровневый анализ бактериальных сообществ, участвующих в колонизации горных пород и почвообразовании в высокогорных условиях. Карр микробиол. 2013; 67: 472–479. 10.1007/s00284-013-0391-9
Трехуровневый анализ бактериальных сообществ, участвующих в колонизации горных пород и почвообразовании в высокогорных условиях. Карр микробиол. 2013; 67: 472–479. 10.1007/s00284-013-0391-9
[PubMed] [CrossRef] [Google Scholar]
2. Wu X, Zhang W, Liu G, Yang X, Hu P, Chen T, et al.
Бактериальное разнообразие в форланде ледника Тянь-Шань № 1, Китай. Environ Res Lett. 2012;7:014038
10.1088/1748-9326/7/1/014038 [CrossRef] [Google Scholar]
3. Sun H, Wu Y, Zhou J, Bing H. Вариации бактериальных и грибковых сообществ вдоль первичной сукцессионной хронопоследовательности в районе отступления ледника Хайлуогоу (гора Гонга). , ЮЗ Китая). J Mt Sci. 2016; 13: 1621–1631. 10.1007/s11629-015-3570-2 [CrossRef] [Google Scholar]
4. Ciccazzo S, Esposito A, Borruso L, Brusetti L. Микробные сообщества и первичная сукцессия в высокогорной среде. Энн Микробиол. 2016; 66: 43–60. 10.1007/s13213-015-1130-1 [CrossRef] [Google Scholar]
5. Тернер Б.Л., Лалиберте Э. Развитие почвы и доступность питательных веществ вдоль 2-миллионной хронологической последовательности прибрежных дюн в богатых видами средиземноморских кустарниках на юго-западе Австралии. Экосистемы. 2015; 18: 287–309. 10.1007/s10021-014-9830-0 [CrossRef] [Google Scholar]
Экосистемы. 2015; 18: 287–309. 10.1007/s10021-014-9830-0 [CrossRef] [Google Scholar]
6. Liu L, Liu Y, Hui R, Xie M. Восстановление структуры микробного сообщества биологических почвенных корок на сукцессионных стадиях восстановления растительности пустыни Шапотоу, северо-запад Китай. Почва Биол Биохим. ООО «Эльзевир»; 2017; 107: 125–128. 10.1016/j.soilbio.2016.12.030 [CrossRef] [Google Scholar]
7. Шауб И., Баум С., Шуманн Р., Карстен У. Влияние ранней сукцессионной биологической почвенной корки из прибрежных песчаных дюн умеренного пояса (СВ Германия) на стехиометрию элементов почвы и активность фосфатазы. Микроб Экол. микробная экология; 2018; 1–13. 10.1007/s00248-018-1220-2
[PubMed] [CrossRef] [Google Scholar]
8. Kelly LC, Cockell CS, Thorsteinsson T, Marteinsson V, Stevenson J. Первые микробные сообщества лавового потока Fimmvortuhals, Eyjafjallajokull, Исландия. Микроб Экол. 2014; 68: 504–518. 10.1007/s00248-014-0432-3 [PubMed] [CrossRef] [Google Scholar]
9. Zhan J, Sun Q. Разнообразие свободноживущих азотфиксирующих микроорганизмов в ризосфере и вне ризосферы пионерных растений, произрастающих на пустырях хвостохранилищ медных рудников. Микробиолог Рез. Исследовательский центр экологических наук Китайской академии наук; 2012; 167: 157–165. 10.1016/j.микр.2011.05.006
Zhan J, Sun Q. Разнообразие свободноживущих азотфиксирующих микроорганизмов в ризосфере и вне ризосферы пионерных растений, произрастающих на пустырях хвостохранилищ медных рудников. Микробиолог Рез. Исследовательский центр экологических наук Китайской академии наук; 2012; 167: 157–165. 10.1016/j.микр.2011.05.006
[PubMed] [CrossRef] [Google Scholar]
10. Cao C, Zhang Y, Cui Z, Feng S, Wang T, Ren Q. Реакция сообщества почвенных бактерий на восстановление растительности движущихся песчаных дюн в полузасушливых пастбищах. Приложение Microbiol Biotechnol. Прикладная микробиология и биотехнология; 2017; 101: 6217–6228. 10.1007/с00253-017-8336-з
[PubMed] [CrossRef] [Академия Google]
11. Yu J, Unc A, Zhang X, Steinberger Y. Реакция микробного катаболического профиля и разнообразия почвы на восстановление растительности на деградированных полузасушливых пастбищах. Прил. Экология почвы. Эльзевир Б.В.; 2016; 101: 124–131. 10.1016/j.apsoil.2016.01.022 [CrossRef] [Google Scholar]
12. Blaud A, Lerch TZ, Phoenix GK, Osborn AM. Микробное разнообразие почв Арктики в меняющемся мире. Рез микробиол. Эльзевир Массон САС; 2015; 166: 796–813. 10.1016/j.resmic.2015.07.013
Blaud A, Lerch TZ, Phoenix GK, Osborn AM. Микробное разнообразие почв Арктики в меняющемся мире. Рез микробиол. Эльзевир Массон САС; 2015; 166: 796–813. 10.1016/j.resmic.2015.07.013
[PubMed] [CrossRef] [Академия Google]
13. Уокер Л.Р., Уордл Д.А., Барджетт Р.Д., Кларксон Б.Д. Использование хронопоследовательностей в исследованиях экологической сукцессии и развития почв. J Экол. 2010; 98: 725–736. 10.1111/j.1365-2745.2010.01664.x [CrossRef] [Google Scholar]
14. Nemergut DR, Anderson SP, Cleveland CC, Martin AP, Miller AE, Seimon A, et al.
Сукцессия микробного сообщества в лишенной растительности недавно дегляциированной почве. Микроб Экол. 2007; 53: 110–122. 10.1007/s00248-006-9144-7
[PubMed] [CrossRef] [Академия Google]
15. Schmidt S., Reed SC, Nemergut DR, Stuart Grandy A, Cleveland CC, Weintraub MN, et al.
Самые ранние стадии сукцессии экосистем в высокогорных (5000 м над уровнем моря), недавно дегляциированных почвах. Proc R Soc B Biol Sci. 2008; 275: 2793–2802. 10.1098/рспб.2008.0808
10.1098/рспб.2008.0808
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Chrismas NAM, Anesio AM, Sánchez-Baracaldo P. Будущее геномики полярных и альпийских цианобактерий. FEMS Microbiol Ecol. 2018; 1–10. 10.1093/фемсек/fiy032
[PMC free article] [PubMed] [CrossRef] [Google Scholar]
17. Glanville HC, Hill PW, MacCarone LD, Golyshin PN, Murphy DV, Jones DL. Температура и вода контролируют появление растительности, динамику микробов и потоки углерода и азота в почве в экосистеме высокогорной арктической тундры. Функция Экол. 2012; 26: 1366–1380. 10.1111/j.1365-2435.2012.02056.x [CrossRef] [Google Scholar]
18. Чапин Ф.С., Матсон П.А., Витоусек П.М. Принципы экологии наземных экосистем [Интернет]. Спрингер;
Нью-Йорк, штат Нью-Йорк: Springer New York; 2011.
10.1007/978-1-4419-9504-9 [CrossRef] [Google Scholar]
19. Castle SC, Sullivan BW, Knelman J, Hood E, Nemergut DR, Schmidt SK, et al.
Питательное ограничение микробной активности почвы на самых ранних стадиях развития экосистемы. Экология. Спрингер Берлин Гейдельберг; 2017; 10.1007/s00442-017-3965-6
Экология. Спрингер Берлин Гейдельберг; 2017; 10.1007/s00442-017-3965-6
[PubMed] [CrossRef] [Google Scholar]
20. Liu X-Y, Koba K, Koyama LA, Hobbie SE, Weiss MS, Inagaki Y, et al.
Нитраты являются важным источником азота для растений арктической тундры. Proc Natl Acad Sci. 2018;115:3398–3403. 10.1073/пнас.1715382115
[Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
21. Henry S, Baudoin E, López-Gutiérrez JC, Martin-Laurent F, Brauman A, Philippot L. Количественная оценка денитрифицирующих бактерий в почвах по гену nirK направленная ПЦР в реальном времени. J Микробиологические методы. 2004; 59: 327–335. 10.1016/j.mimet.2004.07.002
[PubMed] [CrossRef] [Google Scholar]
22. Мэдсен Э.Л. Микроорганизмы и их роль в фундаментальных биогеохимических циклах. Курр Опин Биотехнолог. ООО «Эльзевир»; 2011; 22: 456–464. 10.1016/j.copbio.2011.01.008
[PubMed] [CrossRef] [Академия Google]
23. Браун С.П., Юмппонен А. Сравнение первичных сукцессионных траекторий грибов и бактерий в почвах отступающих ледников. Мол Экол. 2014; 23: 481–497. 10.1111/меч.12487
Мол Экол. 2014; 23: 481–497. 10.1111/меч.12487
[PubMed] [CrossRef] [Google Scholar]
24. Шмидт С.К., Немергут Д.Р., Дарси Дж.Л., Линч Р. По-разному ли собираются сообщества бактерий и грибов во время первичной сукцессии?
Мол Экол. 2014; 23: 254–258. 10.1111/меч.12589
[PubMed] [CrossRef] [Google Scholar]
25. Jangid K, Whitman WB, Condron LM, Turner BL, Williams MA. Сукцессия почвенных бактериальных сообществ в процессе длительного развития экосистемы. Мол Экол. 2013; 22: 3415–3424. 10.1111/меч.12325
[PubMed] [CrossRef] [Академия Google]
26. ИССС. Всемирная справочная база почвенных ресурсов 2014 [Интернет]. 2014. ISSN 0532-0488
27. Фиерер Н., Джексон Дж.А., Вилгалис Р., Джексон Р.Б. Оценка структуры микробного сообщества почвы с использованием таксон-специфических количественных ПЦР-анализов. Appl Environ Microbiol. 2005;71:4117
10.1128/АЭМ.71.7.4117-4120.2005
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
28. Yu Y, Lee C, Hwang S. Анализ структур сообщества в анаэробных процессах с использованием количественного метода ПЦР в реальном времени. Технологии водных наук. 2005; 52: 85–9.1. [PubMed] [Google Scholar]
Анализ структур сообщества в анаэробных процессах с использованием количественного метода ПЦР в реальном времени. Технологии водных наук. 2005; 52: 85–9.1. [PubMed] [Google Scholar]
29. Poly F, Ranjard L, Nazaret S, Gourbière F, Jocteur Monrozier L. Сравнение генофондов nifH в почвах и почвенных микросредах с контрастными свойствами. Appl Environ Microbiol. 2001; 67: 2255–2262. 10.1128/АЭМ.67.5.2255-2262.2001
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
30. Hallin S, Jones CM, Schloter M, Philippot L. Связь между N-циклическими сообществами и функционированием экосистемы в 50-летнем эксперименте по оплодотворению. Исме Дж. Издательская группа Nature; 2009 г.;3: 597–605. 10.1038/Исмей.2008.128
[PubMed] [CrossRef] [Google Scholar]
31. Michotey V, Méjean V, Bonin P. Сравнение методов количественного определения цитохромных cd1-денитрифицирующих бактерий в морских образцах окружающей среды. Appl Environ Microbiol. 2000; 66: 1564–1571. 10.1128/АЭМ.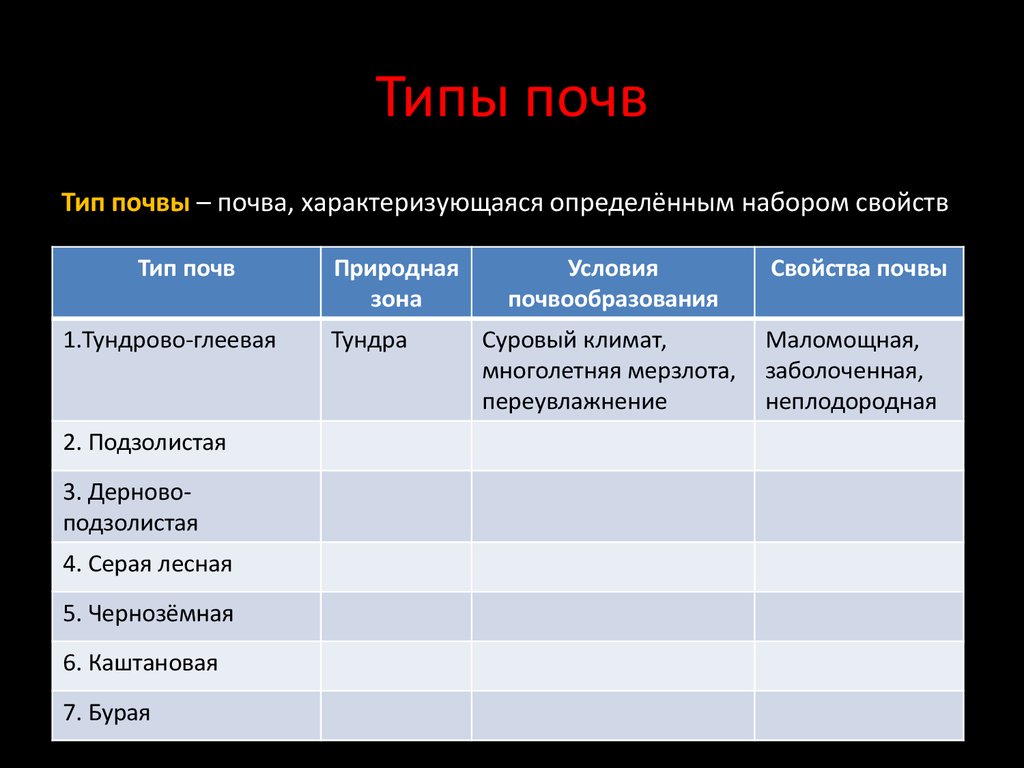 66.4.1564-1571.2000
66.4.1564-1571.2000
[Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
32. Throbäck IN, Enwall K, Jarvis Å, Hallin S. Переоценка праймеров для ПЦР, нацеленных на гены nirS, nirK и nosZ, для общественных исследований денитрифицирующих бактерий с помощью DGGE. FEMS Microbiol Ecol. 2004;49: 401–417. 10.1016/j.femsec.2004.04.011
[PubMed] [CrossRef] [Google Scholar]
33. Bates ST, Berg-Lyons D, Caporaso JG, Walters WA, Knight R, Fierer N. Изучение глобального распределения доминирующих популяций архей в почве. Издательская группа ISME J. Nature; 2011;5: 908–917. 10.1038/исмей.2010.171
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
34. Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al.
QIIME позволяет анализировать данные секвенирования с высокой пропускной способностью. Нат Методы. 2010;7: 335–336. 10.1038/нмет.ф.303
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
35. Bolger AM, Lohse M, Usadel B. Trimmomatic: гибкий триммер для данных последовательностей Illumina. Биоинформатика. 2014;30: 2114–2120. 10.1093/биоинформатика/btu170
Trimmomatic: гибкий триммер для данных последовательностей Illumina. Биоинформатика. 2014;30: 2114–2120. 10.1093/биоинформатика/btu170
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
36. Айва С., Николс Б., Рогнес Т., Флури Т., Маэ Ф. VSEARCH: универсальный инструмент с открытым исходным кодом для метагеномики. Пир Дж. 2016;4: e2584
10.7717/аналог 2584
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Чао А. Непараметрическая оценка количества классов в популяции Автор. Скандинавский J Stat. 1984;11: 265–270. [Google Scholar]
38. Колвелл Р.К., Коддингтон Дж.А. Оценка наземного биоразнообразия посредством экстраполяции. Philos Trans R Soc B Biol Sci. 1994; 345: 101–118. 10.1098/рстб.1994.0091
[PubMed] [CrossRef] [Google Scholar]
39. Lozupone C, Lladser ME, Knights D, Stombaugh J, Knight R. UniFrac: метрика эффективного расстояния для сравнения микробных сообществ. Издательская группа ISME J. Nature; 2011;5: 169–172. 10. 1038/исмей.2010.133
1038/исмей.2010.133
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
40. Бакеридж К.М., Банерджи С., Сицилиано С.Д., Гроган П. Сезонный характер структуры почвенного микробного сообщества в мезональной низинной арктической тундре. Почва Биол Биохим. ООО «Эльзевир»; 2013; 65: 338–347. 10.1016/j.soilbio.2013.06.012 [CrossRef] [Google Scholar]
41. Ren J, Song C, Hou A, Song Y, Zhu X, Cagle GA. Сдвиги в почвенных бактериальных и архейных сообществах во время циклов замерзания-оттаивания в сезонном замерзшем болоте, Северо-Восточный Китай. Научная общая среда. Эльзевир Б.В.; 2018; 625: 782–791. 10.1016/j.scitotenv.2017.12.309[PubMed] [CrossRef] [Google Scholar]
42. Кузяков Ю., Благодатская Е. Микробные очаги и горячие моменты в почве: Концепция и обзор. Почва Биол Биохим. ООО «Эльзевир»; 2015; 83: 184–199. 10.1016/j.soilbio.2015.01.025 [CrossRef] [Google Scholar]
43. Семенов М.В., Чернов Т.И., Тхакахова А.К., Железова А.Д., Иванова Е.А., Колганова Т. В., и др.
В., и др.
Распределение прокариотических сообществ по профилю Черноземья при разных землепользованиях на протяжении более века. Прил. Экология почвы. Эльзевир; 2018; 127: 8–18. 10.1016/j.apsoil.2018.03.002 [CrossRef] [Google Scholar]
44. Regan K, Stempfhuber B, Schloter M, Rasche F, Prati D, Philippot L, et al.
Пространственная и временная динамика азотфиксирующих, нитрифицирующих и денитрифицирующих микробов в неудобренной пастбищной почве. Почва Биол Биохим. ООО «Эльзевир»; 2017; 109: 214–226. 10.1016/j.soilbio.2016.11.011 [CrossRef] [Google Scholar]
45. Ma W, Jiang S, Assemien F, Qin M, Ma B, Xie Z, et al.
Реакция микробных функциональных групп, участвующих в круговороте азота в почве, на удобрения N, P и NP на тибетских альпийских лугах. Почва Биол Биохим. 2016;101:195–206. 10.1016/j.soilbio.2016.07.023 [CrossRef] [Google Scholar]
46. Taş N, Prestat E, Wang S, Wu Y, Ulrich C, Kneafsey T, et al.
Ландшафтная топография структурирует микробиом почвы в арктической полигональной тундре. Нац коммун. 2018;9:777
Нац коммун. 2018;9:777
10.1038/с41467-018-03089-з
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
47. Prosser JI, Nicol GW. Археальные и бактериальные окислители аммиака в почве: поиски специализации и дифференциации ниш. Тенденции микробиол. Текущие тенденции Elsevier; 2012; 20: 523–531. 10.1016/j.tim.2012.08.001
[PubMed] [CrossRef] [Академия Google]
48. Джонс С.М., Халлин С. Экологические и эволюционные факторы, лежащие в основе глобальной и локальной сборки сообществ денитрификаторов. Издательская группа ISME J. Nature; 2010;4: 633–641. 10.1038/исмей.2009.152
[PubMed] [CrossRef] [Google Scholar]
49. Брейкер Г., Фезефельдт А., Витцель К.П. Разработка систем праймеров для ПЦР для амплификации генов нитритредуктазы (nirK и nirS) для обнаружения денитрифицирующих бактерий в пробах окружающей среды. Appl Environ Microbiol. 1998; 64: 3769–75. Доступно: http://aem.asm.org/cgi/content/abstract/64/10/3769.[Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Yang Y, Zhao J, Jiang Y, Hu Y, Zhang M, Zeng Z. Реакция бактерий, несущих гены nirS и nirK, на разную норму азотного удобрения в щелочной среде. Северная китайская земля. Eur J Soil Biol. Эльзевир Массон САС; 2017; 82: 1–9. 10.1016/j.ejsobi.2017.05.006 [CrossRef] [Google Scholar]
Yang Y, Zhao J, Jiang Y, Hu Y, Zhang M, Zeng Z. Реакция бактерий, несущих гены nirS и nirK, на разную норму азотного удобрения в щелочной среде. Северная китайская земля. Eur J Soil Biol. Эльзевир Массон САС; 2017; 82: 1–9. 10.1016/j.ejsobi.2017.05.006 [CrossRef] [Google Scholar]
51. Махаланяне Т.П., Ван Гетхем М.В., Коуэн Д.А. Микробное разнообразие и функциональная способность полярных почв. Курр Опин Биотехнолог. ООО «Эльзевир»; 2016;38:159–166. 10.1016/j.copbio.2016.01.011
[PubMed] [CrossRef] [Google Scholar]
52. Navarrete AA, Tsai SM, Mendes LW, Faust K, De Hollander M, Cassman NA, et al.
Реакция почвенного микробиома на краткосрочные последствия обезлесения Амазонки. Мол Экол. 2015; 24: 2433–2448. 10.1111/меч.13172
[PubMed] [CrossRef] [Google Scholar]
53. Yabe S, Sakai Y, Abe K, Yokota A, Také A, Matsumoto A, et al.
Dictyobacter aurantiacus род. ноябрь, сп. nov., представителя семейства Ktedonobacteraceae, выделенного из почвы, и дополненного описания рода thermosporothrix. Int J Syst Evol Microbiol. 2017; 67: 2615–2621. 10.1099/ijsem.0.001985
Int J Syst Evol Microbiol. 2017; 67: 2615–2621. 10.1099/ijsem.0.001985
[PubMed] [CrossRef] [Google Scholar]
54. Rughoft S, Herrmann M, Lazar CS, Cesarz S, Levick SR, Trumbore SE, et al.
Состав сообщества и обилие бактериальных, архейных и нитрифицирующих популяций в почвах саванны на контрастном материале коренных пород в Национальном парке Крюгера, Южная Африка. Фронт микробиол. 2016;7
10.3389/fmicb.2016.01638
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
55. Li J, Wang JT, Hu HW, Ma YB, Zhang LM, He JZ. Загрязнение медью снижает устойчивость почвенного микробного сообщества к последующему нарушению сухого повторного увлажнения. J Environ Sci (Китай). Эльзевир Б.В.; 2016;39: 155–164. 10.1016/j.jes.2015.10.009
[PubMed] [CrossRef] [Google Scholar]
56. Levy-Booth DJ, Prescott CE, Grayston SJ. Микробные функциональные гены, участвующие в фиксации азота, нитрификации и денитрификации в лесных экосистемах. Почва Биол Биохим. 2014; 75: 11–25. 10. 1016/j.soilbio.2014.03.021 [CrossRef] [Google Scholar]
1016/j.soilbio.2014.03.021 [CrossRef] [Google Scholar]
57. Yarwood SA, Högberg MN. Почвенные бактерии и археи быстро меняются в первом веке освоения фенноскандинавских бореальных лесов. Почва Биол Биохим. 2017; 114: 160–167. 10.1016/j.soilbio.2017.07.017 [CrossRef] [Google Scholar]
58. Bellezza S, Paradossi G, De Philippis R, Albertano P. Штаммы Leptolyngbya из римской гипогеи: цитохимическая и физико-химическая характеристика экзополисахаридов. J Appl Phycol. 2003; 15: 193–200. 10.1023/A:1023811311686 [CrossRef] [Google Scholar]
59. Сиглер В. В., Зейер Дж. Микробное разнообразие и активность на передних полях двух отступающих ледников. Микроб Экол. 2002; 43: 397–407. 10.1007/s00248-001-0045-5
[PubMed] [CrossRef] [Google Scholar]
60. Brankatschk R, Töwe S, Kleineidam K, Schloter M, Zeyer J. Численность и потенциальная активность азотных круговоротов микробных сообществ вдоль хронологической последовательности переднего поля ледника. ISME J. 2011; 5: 1025–1037. 10.1038/исмей.2010.184
10.1038/исмей.2010.184
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
61. Сенгупта А., Дик В.А. Разнообразие метанотрофных бактерий в двух разных почвах при различных методах землепользования, определенное с помощью высокопроизводительного секвенирования гена pmoA. Прил. Экология почвы. Эльзевир; 2017; 119: 35–45. 10.1016/j.apsoil.2017.05.031 [CrossRef] [Google Scholar]
62. Wang S, Zuo X, Zhao X, Awada T, Luo Y, Li Y, et al.
Доминирующие виды растений формируют почвенное бактериальное сообщество в полузасушливых песчаных землях северного Китая. Эколь Эвол. 2018; 8: 1693–1704. 10.1002/экс3.3746
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
63. Сенгупта А., Дик В.А. Разнообразие бактериальных сообществ в почве при двух способах обработки почвы по данным пиросеквенирования. Микроб Экол. 2015; 70: 853–859. 10.1007/s00248-015-0609-4
[PubMed] [CrossRef] [Google Scholar]
Государственный факторный контроль почвообразования во внутренних районах Аляски | Меняющийся бореальный лес Аляски
Фильтр поиска панели навигации
Oxford AcademicИзменяющийся бореальный лес АляскиГеография окружающей средыКнигиЖурналы
Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации
Oxford AcademicИзменяющийся бореальный лес АляскиГеография окружающей средыКнигиЖурналы
Термин поиска на микросайте
Расширенный поиск
Иконка Цитировать
ЦитироватьРазрешения
Делиться
- Твиттер
- Больше
Cite
Packee, Edmond C, and David K Swanson,
‘Государственный факторный контроль почвообразования во внутренних районах Аляски’
,
Alaska’s Changing Boreal Forest
(
New York,
2006;
online edn,
Oxford Academic
, 12 Nov. 2020
2020
), https://doi.org/10.1093 /oso/9780195154313.003.0007,
, по состоянию на 5 декабря 2022 г.
Выберите формат
Выберите format.ris (Mendeley, Papers, Zotero).enw (EndNote).bibtex (BibTex).txt (Medlars, RefWorks)
Закрыть
Фильтр поиска панели навигации
Oxford AcademicИзменяющийся бореальный лес АляскиГеография окружающей средыКнигиЖурналы
Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации
Oxford AcademicИзменяющийся бореальный лес АляскиГеография окружающей средыКнигиЖурналы
Термин поиска на микросайте
Advanced Search
Abstract
Наиболее поразительной особенностью бореальных почв Аляски по сравнению с почвами большинства других биомов является отсутствие значительного развития почвы, хотя большая часть внутренних районов Аляски не была покрыта льдом в течение миллионов лет.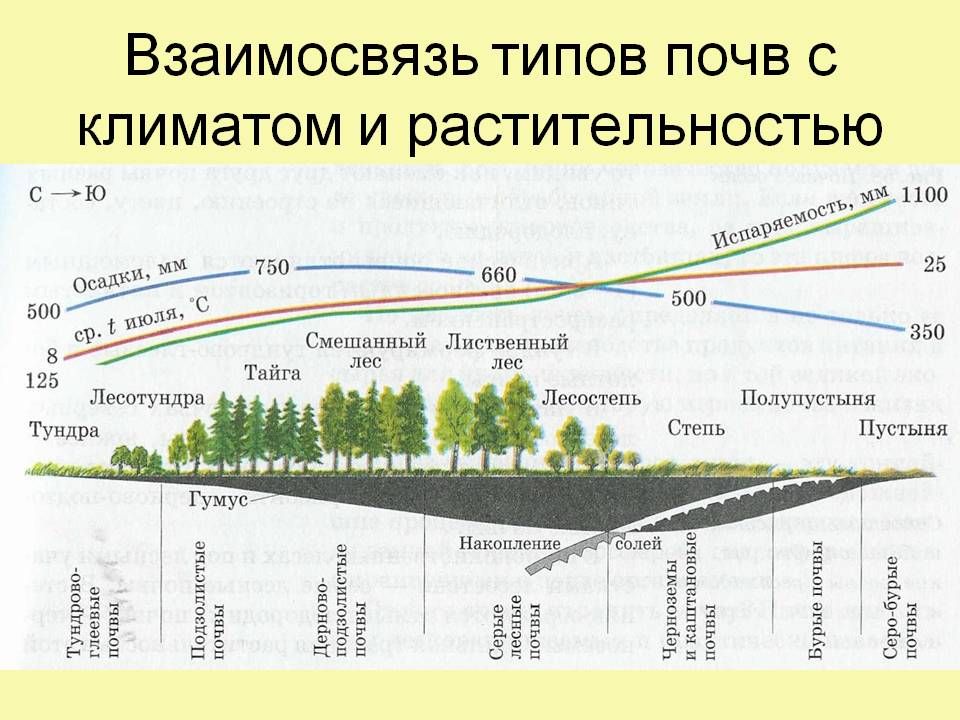 Почвы бореальной зоны поддерживают лесные экосистемы, на долю которых приходится почти половина площади суши Аляски. Во всем мире бореальные леса хранят почти треть наземного углерода (Apps et al., 1993). Следовательно, изменения в бореальных почвах могут сильно повлиять на глобальный углеродный баланс (глава 19).). Еще одна поразительная особенность бореальных почв — их большая локальная изменчивость из-за уклона и рельефа. По сравнению с хорошо развитыми красочными почвами умеренных и тропических регионов, бореальные почвы обычно имеют охристую (желтовато-коричневую) окраску на возвышенностях и темную окраску на низинах из-за накопления органического вещества. На эти низменности приходится 85% запасов водно-болотных угодий в Соединенных Штатах (Бриджем и др., 2001 г.). Почва представляет собой смесь геологического исходного материала и органического вещества, измененного выветриванием и действием живых организмов и обусловленного рельефом с течением времени. Дженни (1941) определил основные факторы, влияющие на почвообразование, такие как исходный материал, организмы, топография, климат и время.
Почвы бореальной зоны поддерживают лесные экосистемы, на долю которых приходится почти половина площади суши Аляски. Во всем мире бореальные леса хранят почти треть наземного углерода (Apps et al., 1993). Следовательно, изменения в бореальных почвах могут сильно повлиять на глобальный углеродный баланс (глава 19).). Еще одна поразительная особенность бореальных почв — их большая локальная изменчивость из-за уклона и рельефа. По сравнению с хорошо развитыми красочными почвами умеренных и тропических регионов, бореальные почвы обычно имеют охристую (желтовато-коричневую) окраску на возвышенностях и темную окраску на низинах из-за накопления органического вещества. На эти низменности приходится 85% запасов водно-болотных угодий в Соединенных Штатах (Бриджем и др., 2001 г.). Почва представляет собой смесь геологического исходного материала и органического вещества, измененного выветриванием и действием живых организмов и обусловленного рельефом с течением времени. Дженни (1941) определил основные факторы, влияющие на почвообразование, такие как исходный материал, организмы, топография, климат и время. Когда изменяется только один из этих факторов, характеристики почвы можно рассматривать как функцию этого фактора (глава 1). Почвы также реагируют на антропогенные изменения, такие как сельское и лесное хозяйство, которые меняют интерактивное управление в местном масштабе (Moore and Ping, 1989). Внутренняя Аляска состоит из нескольких широких, почти ровных низменностей с высотой в основном ниже 500 м и округлых гор с высотой примерно до 2000 м (Wahrhaftig 19).65). Экорегион Внутреннего нагорья (Глава 2) включает Кускоквимское нагорье и Внутреннее нагорье. Экорегион внутренних низин включает низменность Коюкук-Инноко, равнины Канути и низменность Танана-Кускоквим (Rieger et al., 1979). Бассейн реки Коппер к югу от Аляскинского хребта имеет климат и растительность, подобные тем, что на внутренней Аляске. Бассейн реки Коппер имеет мелкозернистые грунты озерного происхождения. Почвы Аляскинского хребта, Кускоквима и Внутреннего нагорья обычно образуются из ледниковых отложений и остаточных материалов и содержат обильные обломки горных пород.
Когда изменяется только один из этих факторов, характеристики почвы можно рассматривать как функцию этого фактора (глава 1). Почвы также реагируют на антропогенные изменения, такие как сельское и лесное хозяйство, которые меняют интерактивное управление в местном масштабе (Moore and Ping, 1989). Внутренняя Аляска состоит из нескольких широких, почти ровных низменностей с высотой в основном ниже 500 м и округлых гор с высотой примерно до 2000 м (Wahrhaftig 19).65). Экорегион Внутреннего нагорья (Глава 2) включает Кускоквимское нагорье и Внутреннее нагорье. Экорегион внутренних низин включает низменность Коюкук-Инноко, равнины Канути и низменность Танана-Кускоквим (Rieger et al., 1979). Бассейн реки Коппер к югу от Аляскинского хребта имеет климат и растительность, подобные тем, что на внутренней Аляске. Бассейн реки Коппер имеет мелкозернистые грунты озерного происхождения. Почвы Аляскинского хребта, Кускоквима и Внутреннего нагорья обычно образуются из ледниковых отложений и остаточных материалов и содержат обильные обломки горных пород.
Ключевые слова:
деятельный слой, насыщенность основания, камбисоль, класс дренированности, экорегион, пойма, гелифлюкция, гистосоль, железо, озерные отложения
Предмет
Экологическая география
В настоящее время у вас нет доступа к этой главе.
Войти
Получить помощь с доступом
Получить помощь с доступом
Доступ для учреждений
Доступ к контенту в Oxford Academic часто предоставляется посредством институциональных подписок и покупок. Если вы являетесь членом учреждения с активной учетной записью, вы можете получить доступ к контенту одним из следующих способов:
Доступ на основе IP
Как правило, доступ предоставляется через институциональную сеть к диапазону IP-адресов. Эта аутентификация происходит автоматически, и невозможно выйти из учетной записи с IP-аутентификацией.
Эта аутентификация происходит автоматически, и невозможно выйти из учетной записи с IP-аутентификацией.
Войдите через свое учреждение
Выберите этот вариант, чтобы получить удаленный доступ за пределами вашего учреждения. Технология Shibboleth/Open Athens используется для обеспечения единого входа между веб-сайтом вашего учебного заведения и Oxford Academic.
- Щелкните Войти через свое учреждение.
- Выберите свое учреждение из предоставленного списка, после чего вы перейдете на веб-сайт вашего учреждения для входа.
- Находясь на сайте учреждения, используйте учетные данные, предоставленные вашим учреждением. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если вашего учреждения нет в списке или вы не можете войти на веб-сайт своего учреждения, обратитесь к своему библиотекарю или администратору.
Вход с помощью читательского билета
Введите номер своего читательского билета, чтобы войти в систему. Если вы не можете войти в систему, обратитесь к своему библиотекарю.
Члены общества
Доступ члена общества к журналу достигается одним из следующих способов:
Войти через сайт сообщества
Многие общества предлагают единый вход между веб-сайтом общества и Oxford Academic. Если вы видите «Войти через сайт сообщества» на панели входа в журнале:
- Щелкните Войти через сайт сообщества.
- При посещении сайта общества используйте учетные данные, предоставленные этим обществом. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если у вас нет учетной записи сообщества или вы забыли свое имя пользователя или пароль, обратитесь в свое общество.
Вход через личный кабинет
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам. Смотри ниже.
Личный кабинет
Личную учетную запись можно использовать для получения оповещений по электронной почте, сохранения результатов поиска, покупки контента и активации подписок.
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам.
Просмотр учетных записей, вошедших в систему
Щелкните значок учетной записи в правом верхнем углу, чтобы:
- Просмотр вашей личной учетной записи и доступ к функциям управления учетной записью.
- Просмотр институциональных учетных записей, предоставляющих доступ.
Выполнен вход, но нет доступа к содержимому
Oxford Academic предлагает широкий ассортимент продукции.